
浙江近海春、夏季蟹类群落结构及其与环境因子的关系
2019-05-29 06:58:42卢衎尔张洪亮朱文斌徐开达周永东卢占晖
水生生物学报 2019年3期
卢衎尔 张洪亮 朱文斌 徐开达 周永东 卢占晖
(浙江省海洋水产研究所, 浙江省海洋渔业资源可持续利用技术研究重点实验室, 农业部重点渔场渔业资源科学观测实验站, 舟山 316004)
浙江沿岸海域受长江冲淡水、台湾暖流、苏北沿岸水和黄海冷水团等海流和水系的综合影响,加之沿岸海域岛屿分布众多, 各水系交流充分, 营养盐类和饵料生物丰富, 是多种包括蟹类在内的渔业生物的产卵场和索饵场[1]。目前, 国外对于近岸和大陆架海域蟹类生态学方面的研究日益活跃, 如Jason等[2]研究了美国新罕布什尔州Great Bay Estuary海域Carcinus maenas的分布和丰度, Bertini等[3]对巴西圣保罗州北部近岸海域蟹类生物多样性进行了探讨, Andrade等[4]研究了巴西东南部大陆架海域梭子蟹(Portunoidea)的群聚组成和结构。我国早期对于海洋蟹类方面的研究, 主要集中于种类组成与区系特征方面[5,6]。20世纪90年以来, 对于东海范围内蟹类资源分布[7,8]和群落生态学[9—11]方面的研究也被陆续报道, 但这些报道的研究区域大部分集中在30 m等深线以深海域, 且调查时间主要集中在1998年或2008年, 难以准确反映当前蟹类群落的现状。有关东海近岸水域蟹类群落方面的研究则集中在典型局部(如河口)海域[12—14], 涉及的研究区域普遍较小。
本研究基于2015年浙江近岸海域渔业资源大面定点调查数据, 研究分析了蟹类群落的种类组成及区系特征、优势种组成、资源密度分布及群落物种多样性等内容, 并首次采用典范对应分析(CCA)方法阐述了蟹类群落结构与海洋环境因子之间的关系。此研究的开展, 有助于丰富中国近海海域蟹类生物多样性的基础资料, 对于认识浙江乃至整个东海海域蟹类最新的资源动态变化有一定的科学意义, 同时也为浙江近海海洋生态和环境保护措施的制定提供参考依据。
1 材料与方法
1.1 数据来源
本文所用数据来自2015年4月(春季)和7月(夏季)进行的“浙江沿岸产卵场调查与选划”课题。共设置调查站位87个, 站位分布如图1所示。调查船为单船底拖网渔船, 船长38 m, 总吨位150 t, 主机功率202 kW。调查网具网口拉紧周长50 m, 囊网网目尺寸25 mm, 上纲长30 m, 下纲长38 m。各站位拖网时间均为1h, 平均拖速3 kn。水深、温度、盐度、溶解氧和叶绿素a等相关环境因子数据采用多功能水质参数仪与拖网调查同步采集, 每个站位各1次。各站位渔获物样品随机采样1箱, 渔获物中蟹类种类尽可能鉴定到最小分类单元, 并记录每种的生物量和尾数, 蟹类种类名称、分类地位以及适温类型以《中国海洋生物名录》[15]为依据, 同时参照《东海经济虾蟹类》[16]和Sealifebase数据库(http://www.sealifebase.org)。上述调查采样及测定均按《海洋调查规范(GB/T 12763.6—2007)》[17]的有关标准进行。

图1 浙江近岸海域调查站位分布图Fig. 1 Location of sampling sites in the Zhejiang coast area
1.2 分析方法
优势种与常见种群落优势种与常见种采用Pinkas的相对重要性指数(Index of relative importance,IRI)进行划分[18]。其公式为:

式中,N%为某一种类的尾数占总尾数的百分比;W%为某一种类的生物量占总生物量的百分比;F为某一种类出现的站数占调查总站数的百分比。
资源密度蟹类主要营底栖生活, 因此, 本研究采用扫海面积法对调查海域蟹类资源密度分布进行分析。其计算公式为:
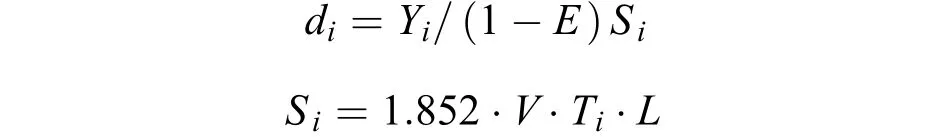
式中,di为i站位的资源密度(kg/km2),Yi为调查船在i站位的渔获量(kg);Si为调查船在i站位的扫海面积(km2);E为逃逸率(根据本次调查网具——单船底拖网的结构, 渔法特点以及蟹类分布水层和生活习性,综合相关研究报告逃逸率E的取值依据[7,19], 本研究选取的逃逸率为0.7);V为网具拖曳的平均拖速(kn)(本次调查拖网拖曳平均拖速为3 kn);Ti为拖网时间(本次调查均为1h);L为网口水平扩张宽度(km)(本次调查网具为5×10-3km)。
群落多样性本文蟹类群落多样性分析运用下列公式[20], 种类丰富度指数D:
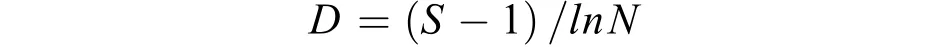
Shannon-Wiener多样度指数H′:
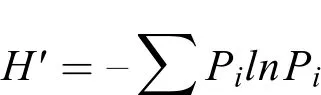
种类均匀度指数J′:

式中,S为种类数;N为总尾数;W为总生物量;Pi为i种蟹类占总生物量的比例。
群落ABC曲线Warwick1986年提出的丰度/生物量比较曲线(Abundance/Biomass curves,ABC曲线) 通过丰度和生物量优势度曲线比较来监测干扰对底栖无脊椎动物群落的影响。本文根据ABC曲线中生物量和丰度的K-优势度曲线, 分析检验浙江近海海域蟹类群落受到污染和扰动的情况。根据Warwick和Clarke的理论, 若生物量优势度曲线位于丰度优势度曲线之上, 则表明群落处于未受干扰(稳定)的状态; 若两条曲线相交, 则表明群落处于中等干扰(或不稳定)的状态; 若生物量的优势度曲线在丰度优势度曲线之下, 则表明群落处于严重干扰的(不稳定的)状态。用W统计量(W-statistic)作为ABC曲线方法的一个统计量, 其公式为式中Bi和Ai为ABC曲线中种类序号对应的生物量和丰度的累积百分比,S为出现物种数。当生物量优势曲线在丰度优势度曲线之上时,W为正, 反之W为负[21]。
群落与环境因子的关系典范对应分析(Canonical Correspondence Analysis, CCA)是一种非线性多元直接梯度分析方法[22], 其特点是可以结合多个环境因子一起分析, 包含的信息量大, 结果直观明显, 从而更好地反映群落与环境的关系[23]。对浙江近岸蟹类群落与水深、温度、盐度、溶解氧和叶绿素a(均为表底层)等9个环境变量进行CCA排序, 通过蒙特-卡洛置换检验从中筛选出对群落有显著影响的环境变量, 并做出蟹类群落空间分布与环境因子关系的二维排序图。
在以上数据分析中, 群落的物种多样性指数、ABC曲线及W统计量的计算均使用PRIMER6.0软件进行处理; 典范对应分析(CCA)在CANOCO4.5软件进行运算, 排序结果用物种-环境因子关系的双序图表示, 站位图、资源密度和多样性指数分布图利用surfer作图软件进行绘制。
2 结果
2.1 海洋环境因子
浙江近岸春、夏季各站位环境因子参数平均值见表1。数据分析表明: 各站位的平均水深为37.78 m, 叶绿素a(表层、底层)、溶解氧(表层、底层)和底层盐度等5个环境因子参数的季节变化一致, 均呈现春季高于夏季的趋势, 温度(表层、底层)和表层盐度等3个环境因子参数呈现夏季高于春季的趋势。

表1 春、夏季浙江近海海洋环境因子参数(均值±标准误)Tab. 1 Environmental factors of 87 samples in the Zhejiang coast area in spring and summer (Mean±SE)
2.2 种类组成与分布
春、夏季两次调查中共采集蟹类28种, 隶属于11科, 19属(表2)。其中, 梭子蟹科出现种类数最多,有11种, 占蟹类总种类数的39.29%; 经济蟹类有三疣梭子蟹(Portunus trituberculatus)、锯缘青蟹(Scylla serrata)、日本蟳(Charybdis japonica)、细点圆趾蟹(Ovalipes punctatus)、中华绒螯蟹(Eriochier sinensis)、锈斑蟳(Charybdis feriatus)、红星梭子蟹(Portunus sanguinolentus)和武士蟳(Charybdis miles)等8种, 其余均为非经济种类或小型蟹类;从不同种类的适温类型上看, 调查海域蟹类由暖水性和暖温性两种类型组成, 暖水性占绝对优势, 种类16数达22种, 占总种类数的78.57%, 暖温性种类6种, 占21.43%。从种类组成的季节分布看, 春季和夏季两个出现蟹类种类均为22种, 春季种类隶属于8科, 14属; 夏季种类隶属于10科, 16属; 种类组成呈现一定的季节变化, 春夏两季均出现的种类有16种,占总种类数的57.14%, 春季和夏季特有种均为6种。
春季不同站位间出现的蟹类种类数最多的为6种, 出现在调查海域北部外侧区域的33和34号站位, 最少的仅有1种, 出现在27、42、49、71、72和82号站位, 春季不同站位出现蟹类种类分布较为均匀, 仅在调查海域中北部出现种类数相对较多, 空间分布特征不明显; 夏季不同站位间出现的蟹类种类数最多的为9种, 出现在调查海域北部外侧区域的4号站位, 最少的仅有1种, 出现在调查海域内侧区域的8、30和65号站位, 夏季蟹类种类的空间分布特征较春季明显, 呈现中间低、两侧高的特征(图2)。
2.3 优势种与常见种
利用相对重要性指数(IRI)来确定各种类在虾类群落中的重要性。本文中将相对重要性指数(IRI)大于500的定为优势种,IRI值在50—500的种类定为常见种(表3)。根据上述规定, 春季蟹类群落的优势种按IRI值大小依次为双斑蟳、三疣梭子蟹和日本蟳, 这三种蟹类渔获质量占该航次蟹类总渔获物质量的84.78%, 渔获尾数占蟹类总渔获物尾数的92.94%, 常见种为细点圆趾蟹和长手隆背蟹;夏季蟹类群落的优势种按IRI值大小依次为三疣梭子蟹和双斑蟳, 这两种蟹类渔获质量占该航次蟹类总渔获物质量的89.96%, 渔获尾数占蟹类总渔获物尾数的94.82%, 常见种为细点圆趾蟹和日本蟳。总体来看: 调查海域春、夏季蟹类群落的优势种与常见种种类组成基本相同, 且三疣梭子蟹和双斑蟳为两个季节共同的绝对优势种。
2.4 资源密度
春、夏两个航次调查中共采集蟹类140355 ind.,生物量996.84 kg, 平均个体生物量7.10 g, 2个航次
调查总站位数为174个, 其中出现蟹类的站位数有174个, 出现频率100%。夏季蟹类平均资源密度明显高于春季。

表2 春、夏季浙江近海蟹类群落种类名录Tab. 2 List of crab species in the Zhejiang coast area in spring and summer

图2 浙江近岸海域春、夏季蟹类种类空间分布(a. 春季; b. 夏季)Fig. 2 The space distribution of crabs in the Zhejiang coast area in spring and summer (a. spring; b. summer)
春季: 蟹类平均资源密度为244.38 kg/km2, 资源密度最小值为1.19 kg/km2, 出现在位于鱼山渔场的72号站位, 资源密度最大值为2043.85 kg/km2, 出现在位于舟山渔场的2号站位; 根据空间分布状况(图3), 整个调查海域蟹类资源密度分布较为均匀,但29°00′N以北的调查海域资源密度较南部海域略高。
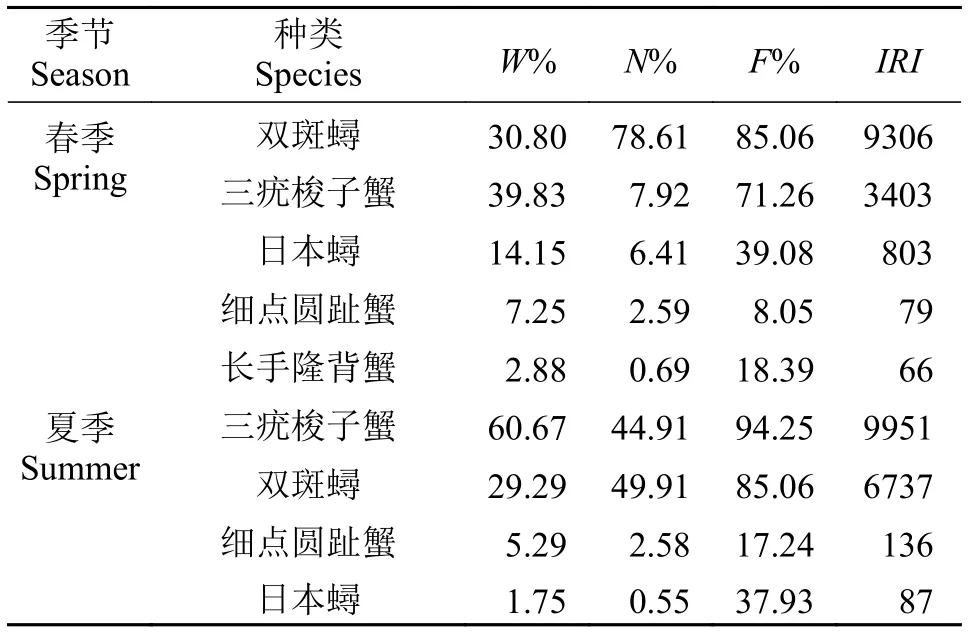
表3 春、夏季浙江近海蟹类群落优势种与常见种组成Tab. 3 The dominant and common species of shrimp community in the Zhejiang coast area in spring and summer
夏季: 蟹类平均资源密度为585.60 kg/km2, 资源密度最小值为4.71 kg/km2, 出现在位于鱼山渔场的65号站位, 资源密度最大值为3955.04 kg/km2, 出现在位于鱼山渔场的49号站位; 根据空间分布状况(图2), 浙江中北部近岸海域, 即28°30′N以北的调查区域, 蟹类平均资源密度明显高于浙南海域。

图3 浙江近岸海域春、夏季蟹类资源密度分布(a. 春季; b. 夏季)Fig. 3 The density distribution of crabs in the Zhejiang coast area in spring and summer (a. spring; b. summer)
2.5 物种多样性
选取3个具有代表性的生物多样性特征值: 种类丰富度指数(D)、以生物量计算的Shannon-Wiener多样度指数(H′)和以生物量计算的种类均匀度指数(J′)来分析浙江近岸海域春、夏季蟹类群落多样性特点(表4)。经计算可得: 春季, 种类丰富度指数(D)、物种多样性指数(H′)和种类均匀度指数(J′)的平均值分别为0.27、0.62和0.58, 夏季3个多样性指数的值依次为0.27、0.61和0.54。春、夏季3种多样性指数数值均较小, 蟹类群落多样性水平处于较低水平。单因素方差分析表明, 浙江近岸海域蟹类群落结构多样性指数春、夏季差异不显著(P>0.05)。春、夏两个季节调查海域蟹类3个多样性指数的空间分布特征趋于一致: 均呈现由西至东逐渐升高的趋势, 即调查区域的近岸区域多样性指数低于外海区域(图4)。

图4 浙江近岸海域春、夏季蟹类群落生物多样性指数分布Fig. 4 The biodiversity indexes distribution of crabs community in the Zhejiang coast area in spring and summer
2.6 ABC曲线
如图5所示, 春季调查海域蟹类的丰度优势度曲线位于生物量曲线之上, 夏季的丰度优势度曲线与生物量的优势度曲线相交。2个季节根据ABC曲线计算所得的W统计值分别为-0.069和0。根据Warwick和Clarke的理论, 春季蟹类群落处于严重干扰状态, 而夏季则处于中等干扰(或不稳定)的状态[21]。

图5 春、夏季浙江近岸海域蟹类群落的ABC曲线以及W统计值(▽. 生物量; △. 丰度)Fig. 5 ABC curves and W value of crabs community the Zhejiang coast area in spring and summer (▽. Biomass; △. Abundance)
2.7 群落与环境因子关系的CCA分析
采用站位-种类生物量矩阵和站位-环境因子矩阵对浙江近岸蟹类群落进行CCA排序, 结果表明:物种CCA排序轴第一轴和第二轴与环境因子极显著相关, 其特征值(Eigenvalues)分别为0.406、0.172(春季)和0.331、0.190 (夏季), 前两个环境因子排序轴与种类排序轴之间的相关系数分别为87.5%、69.9% (春季)和85.6%、73.5% (夏季), 前两个种类排序轴近似垂直(相关系数-0.0914 (春季)和-0.0131(夏季)), 前两个环境排序轴的相关系数均为0, 两个季节CCA排序轴前四轴种类-环境方差累计贡献率分别达到81.9%和86.2%, 蒙特卡罗拟合检验表明第一轴和前四轴所代表的环境变量均与种类变量之间呈显著相关关系(春季: 第一轴F=7.045,P=0.002,前四轴F=2.132,P=0.002; 夏季: 第一轴F=8.386,P=0.002, 前四轴F=2.926,P=0.002)。以上均证实排序轴与环境因子间线性结合的程度较好地反映了群落与环境之间的关系, 排序的结果是可靠的[22]。本研究采用前两轴绘制种类-环境因子的二维空间排序图(图6)。
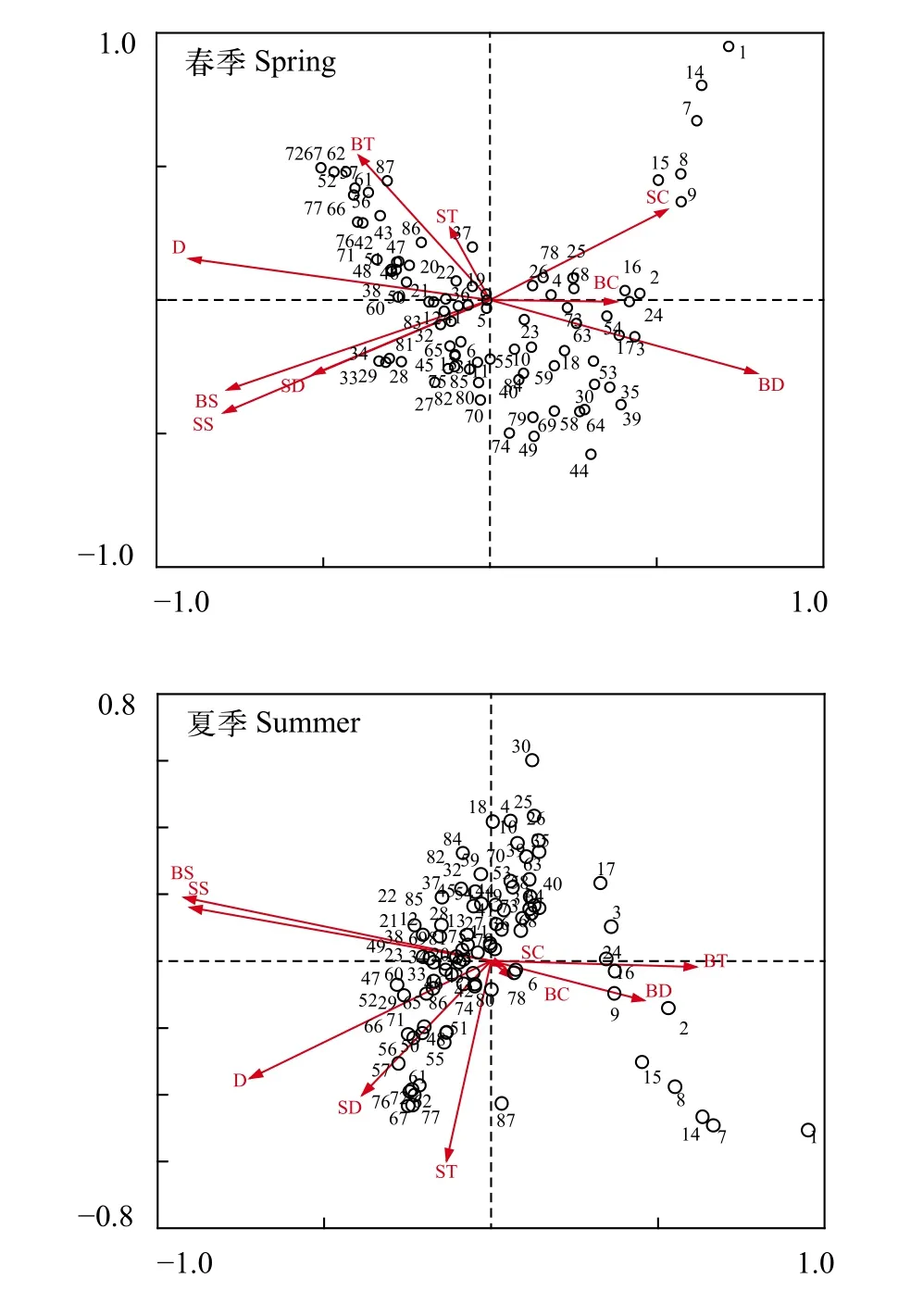
图6 春、夏季蟹类群落不同调查站位的CCA二维排序图Fig. 6 CCA ordination biplot between different sampling stations and environmental factors in spring and summer
综合调查站位的CCA二维排序图(图6)和环境因子与排序轴的相关性大小(表5)可以得出, 春季,CCA排序第一轴主要反映了以下多个环境因子的综合变化趋势: 与底层溶解氧呈极显著正相关, 相关系数为0.7001; 与水深、表层盐度和底层盐度呈极显著负相关, 相关系数分别为-0.7890、-0.6981和-0.6890, 即沿CCA排序轴第一轴从左到右底层溶解氧逐渐升高, 水深和盐度(表层和底层)逐渐降低, 在所有9个环境因子中, 水深与第一轴的相关性最大。CCA排序第二轴与9个环境因子之间均无极显著相关, 其中与底层温度相关性最大, 相关系数为0.3775, 即沿CCA排序轴第二轴从下到上, 底层温度逐渐升高。综合前两轴相关性结果: 水深、底层溶解氧和盐度(表层和底层)等4个环境因子对春季蟹类群落的分布起较大的作用。

表5 春、夏季蟹类群落环境因子与CCA环境轴的相关性Tab. 5 Correlation between environmental factors of crabs communities and CCA environmental axis in spring and summer
夏季, CCA排序第一轴主要反映了以下多个环境因子的综合变化趋势: 与底层温度呈极显著正相关, 相关系数为0.5255; 与底层盐度、表层盐度和水深呈极显著负相关, 相关系数分别为-0.7870、-0.7707和-0.6168, 即沿CCA排序轴第一轴从左到右底层温度逐渐升高, 盐度(表层和底层)和水深逐渐降低, 在所有9个环境因子中, 底层盐度与第一轴的相关性最大。CCA排序第二轴与9个环境因子之间均无极显著相关, 其中与表层温度相关性最大,相关系数为-0.4389, 即沿CCA排序轴第二轴从下到上, 表层温度逐渐降低。综合前两轴相关性结果: 盐度(表层和底层)、水深和底层温度等4个环境因子对夏季蟹类群落的分布起较大的作用。
3 讨论
3.1 蟹类群落结构特征分析
在本次调查中共捕获蟹类种类28种, 隶属于11科, 19属, 其种类数远低于俞存根等[9]的研究结果(45种)。这主要是由于本次调查只在春、夏两季进行, 加之历史调查中的站位绝大部分集中在拖网禁渔区线至127°E海域, 能够捕获更多的外海种类。与瓯江口、浙江南部近岸、中街山列岛以及马鞍列岛[12,14,24,25]等浙江沿岸局部海域调查出现的蟹类种类(种类数范围8—22种)相比, 本次调查基本涵盖了浙江近海蟹类的主要种类。同其他相邻海区相比, 本次调查出现蟹类种类数高于黄海山东近岸海域[26], 而明显低于南海北部近岸种类数[27], 这也能够从一定程度上说明热带海域较亚热带和温带海域具有更多蟹类种类。浙江近岸春夏季蟹类种类的区系成分较简单, 以暖水种占优势, 无冷水性种类出现, 基本属于印度洋-西太平洋动物区系范畴,这与沈嘉瑞等[6]的研究结论一致。浙江近海春、夏两季蟹类群落共出现3种优势种, 分别为双斑蟳、三疣梭子蟹和日本蟳, 以上3种蟹类均属于广温广盐类群, 这也说明浙江近岸海域蟹类群落以该种类群占主导地位。三疣梭子蟹和双斑蟳是两个季节的共有优势种, 且优势度明显高于日本蟳以及群落中的其他蟹类种类, 说明研究海域蟹类优势种较为稳定。与浙江沿岸历史数据[12,14,24,25]相比较, 优势种的种类组成也基本无变化, 唯一的不同就是, 三疣梭子蟹的优势度呈现较大幅度的提升, 究其原因: 一方面是由于三疣梭子蟹在春夏季在浙江沿岸海域进行产卵与索饵[16], 在此期间更容易被捕获;另一方面也可能与近年来浙江近海三疣梭子蟹大规模增殖放流从而提升了群体数量有关。根据调查结果, 浙江近岸春夏两季群落多样性指数季节间变化不明显, 且整体水平偏低, 群落多样性水平不高, 群落处于相对不稳定状态。3个群落多样性水平指数的空间分布特征均呈现沿岸低, 外海高的趋势。其中, 尤以物种多样性指数(H′)空间分布特征最为明显, 这说明调查海域蟹类种类数和分布的均匀程度自西向东逐渐提高。这与俞存根等[9]在整个东海海域的研究结果一致。
3.2 资源密度分布特征
调查海域春夏两季蟹类平均资源密度为414.99 kg/km2, 夏季蟹类平均资源密度明显高于春季, 主要是由于春季蟹类群体中主要以产卵群体为主, 群体数量较小, 而夏季蟹类群体主要以当年生的蟹类补充群体为主, 规模和数量都较春季大幅度增加, 这一现象与俞存根等[7]在东海大陆架海域的研究结果一致。根据图2所体现的资源密度空间分布趋势可得: 浙江近岸海域蟹类资源密度呈现由北向南逐渐降低的趋势, 这主要是因为作为影响资源密度分布的双斑蟳和三疣梭子蟹在春、夏季主要分布在浙江中北部近岸海域。
3.3 扰动对蟹类群落的影响
综合调查海域春、夏季蟹类群落ABC曲线特征及W统计值, 根据Clarke和Warwick的划分标准,表明调查海域春季蟹类群落处于严重干扰状态, 而夏季蟹类群落较春季稳定, 处于中度干扰状态。究其原因, 主要是因为随着浙江近岸捕捞力量的不断增强, 捕捞行为已经成为影响研究海域蟹类群落稳定最为重要的因素之一, 而春季(4月份)调查期间正处于浙江渔场的捕捞汛期, 人类捕捞活动频繁, 使得春季蟹类群落处于严重干扰状态; 而夏季(7月份)则恰好处于东海区“伏季休渔”期间, 以蟹类为主要渔获对象的蟹笼、刺网、桁杆拖虾等海洋捕捞作业方式全部处于休渔期, 人为扰动对蟹类群落的影响降到了最低, 加之蟹类群落的优势种之一三疣梭子蟹的增殖放流活动主要集中在夏季, 提高了研究海域蟹类群落的多样性水平, 以上两类因素使得夏季的蟹类群落较之春季更加稳定。这与李圣法[28]在评价东海鱼类群落稳定性中的研究结论相吻合。
3.4 蟹类群落与环境因子的关系
本研究对浙江近海蟹类与环境因子进行CCA分析, 综合两个季节不同环境因子与站位分布的相关性大小以及环境因子的分布特征可得: 在进行对应分析的9个环境因子中, 水深在春夏季与蟹类群落的分布均呈极显著负相关, 且相关系数较大, 为影响研究海域春夏两季蟹类分布的首要环境因子,这主要由于这两个季节为包括三疣梭子蟹[29]和双斑蟳[30]在内的蟹类的产卵期和幼体索饵发育期, 而无论是产卵亲体还是补充群体(幼体)均趋向于水深较浅的海域栖息分布, 资源密度的分布特征(图2)同样也验证了这一规律。这与杨刚等[31]在黄海蟹类群落分布中的研究结论一致, 也同样体现在海洋鱼类群落格局的分布规律上[32]; 盐度(表层和底层)为影响蟹类分布的次要环境因子, 但夏季的相关系数较春季略高, 这主要是由于夏季较春季降雨增加, 长江冲淡水径流量增大, 导致研究海域盐度梯度变化加大。根据表1也可以看出: 本研究实测结果中夏季表层盐度和底层盐度标准误(SE)远高于春季, 同样也印证了以上的表述。这也与徐兆礼[12]的研究结果一致。底层溶解氧和底层温度在春季和夏季交替成为影响群落分布的环境因子, 其主要是通过水深的变化从而影响以上两种环境因子的变化。此外, 需要指出的是: 蟹类群落结构受多种环境因子综合作用的影响, 但由于调查项目的局限性, 诸如营养盐、悬浮物、pH、沉积物粒径、浮游动植物丰度等生物和非生物环境因子对蟹类分布与群落结构的影响未能涉及, 这也有待于进一步的研究。
猜你喜欢
热带林业(2022年4期)2023-01-10 10:44:04
今日农业(2021年8期)2021-11-28 05:07:50
华东师范大学学报(自然科学版)(2021年2期)2021-11-10 03:10:35
水上消防(2021年3期)2021-08-21 03:12:00
江苏农业科学(2021年3期)2021-04-29 16:12:13
人大建设(2019年5期)2019-10-08 08:55:10
收藏界(2018年1期)2018-10-10 05:23:08
创新作文(小学版)(2018年31期)2018-05-16 03:00:30
摄影之友(影像视觉)(2017年1期)2017-07-18 11:12:16
安徽农业科学(2017年26期)2017-06-11 16:38:25
