
高糖环境下IQGAP1在足细胞中的表达及其对足细胞骨架重排的调节作用▲
2019-05-22 08:29:00陈星华杨英杰李紫燃
广西医学 2019年7期
黄 静 张 璐 陈星华 杨英杰 李紫燃 程 晖
(武汉大学人民医院肾内科,湖北省武汉市 430000,电子邮箱:huang.jingkl@163.com)
糖尿病肾病是引起终末期肾衰竭的主要病因之一。肾小球脏层上皮细胞即足细胞,是肾小球滤过膜的最后一道屏障,其损伤对糖尿病肾病蛋白尿的发生与进展的影响日益受到重视[1]。足突融合是足细胞损伤的特征性微观病理改变,其与足细胞骨架重排密切相关[2]。在正常足突中,细胞骨架蛋白纤维状肌动蛋白(fibrous actin,F-actin)有序地平行聚集成束,形成应力纤维。病理状态下,骨架发生重排,F-actin解聚或以松散的网状机构互相结合。骨架重排是足突融合的共同通路[2],是糖尿病肾病发生的关键环节之一。然而糖尿病肾病足细胞骨架重排的机制尚未明确。
IQ结构域GTP酶活化蛋白1(IQ domain GTPase-activating protein 1,IQGAP1)是一个含有多个蛋白结合域的支架蛋白。Schwarz等[3]的研究发现,IQGAP1蛋白在全身多个组织和细胞中表达,并在足细胞中大量表达。本课题组的前期研究显示IQGAP1在嘌呤霉素诱导的足细胞骨架重排中发挥重要作用[4]。但IQGAP1是否参与糖尿病肾病的足细胞骨架重排尚未见报道。本课题旨在研究IQGAP1在高糖诱导的足细胞骨架重排中的作用及其可能的机制,进一步探讨糖尿病肾病条件下足细胞损伤的分子机制。
1 材料与方法
1.1 主要试剂 胎牛血清(美国Gibco公司,批号:1900-141),1640培养基(武汉普诺赛生命科技有限公司,批号:WH01111810XP),小鼠重组干扰素(美国PeproTech公司,批号:315-05),细胞裂解液(上海碧云天生物技术有限公司,批号:P0013),IQGAP1兔多克隆抗体(美国Santa Cruz公司,批号:sc-10792),细胞分裂周期蛋白42(cell division cycle 42,Cdc42)兔多克隆抗体(美国Cell Signaling公司,批号:2462S),β-肌动蛋白(β-actin)鼠单克隆抗体(美国Santa Cruz公司,批号:YSm-33036M),鬼笔环肽(美国Sigma公司,批号:P5282),X-tremeGENE转染试剂(Roche公司,批号:6366244001),免疫沉淀试剂盒(上海碧云天生物技术有限公司,批号:P2012)。
1.2 细胞培养及分组 永生化小鼠足细胞株由美国纽约Peter Mundel教授惠赠。常规复苏细胞后,用含10%胎牛血清、10 U/mL小鼠重组干扰素的1640培养基,置于33℃、5%CO2培养箱中进行增殖培养7~10 d,然后用含10%胎牛血清的无干扰素1640培养基,置于37℃、5% CO2培养箱中分化培养10~14 d后用于后续实验。细胞分组:(1)将细胞分为6组,分别加入不同浓度葡萄糖(5 mmol/L、10 mmol/L、15 mmol/L、20 mmol/L、25 mmol/L、30 mmol/L)刺激48 h,然后检测各组IQGAP1蛋白的表达情况;(2)另取细胞分为6组,加入高浓度葡萄糖(30 mmol/L)刺激不同时间(0 h、3 h、6 h、12 h、24 h、48 h),然后检测各组IQGAP1蛋白的表达情况;(3)另取细胞分为6组,包括正常糖浓度组(5 mmol/L葡萄糖,NG组)、高糖组(30 mmol/L葡萄糖,HG组)、高渗对照组(5 mmol/L葡萄糖+25 mmol/L甘露醇,HO组)、HG+IQGAP1质粒转染组、HG+空质粒转染组、NG+IQGAP1质粒转染组、NG+空质粒转染组,每组均经葡萄糖刺激48 h,质粒转染方法详见1.3,干预后检测各组IQGAP1蛋白或Cdc42蛋白表达情况。
1.3 质粒转染IQGAP1 IQGAP1全长表达载体(pCantag-myc-IQGAP1质粒)由美国康奈尔大学Jon Erickson教授惠赠。取足细胞分别应用5 mmol/L葡萄糖、30 mmol/L葡萄糖刺激48 h后,将正常糖浓度组及高糖组足细胞接种于6孔板中(25 cm2/孔)培养至足细胞融合50%左右时进行转染,将2 μg质粒与6 μL转染试剂混匀,加入200 μL无血清1640培养基中,30 min后加入培养细胞,37℃恒温培养箱中孵育48~72 h。空质粒转染方法同上。
1.4 免疫印迹法检测IQGAP1和Cdc42蛋白表达 细胞裂解液裂解NG组、NG+IQGAP1质粒转染组、NG+空质粒转染组、HG组、HG+IQGAP1质粒转染组、HG+空质粒转染组细胞,4℃ 2 500 r/min低温离心5 min,后取上清,二喹啉甲酸BCA法测定蛋白浓度,100℃变性5 min,以20 μg/孔上样,10%十二烷基硫酸钠(sodium dodecyl sulfate,SDS)-聚丙烯酰胺凝胶(polyacrylamide gel electrophoresis,PAGE)电泳,湿转至聚偏二氟乙烯膜,5%脱脂牛奶37℃封闭1 h,一抗分别为IQGAP1兔多克隆抗体(1 ∶200)、Cdc42兔多克隆抗体(1 ∶1 000),4℃孵育过夜,β-actin鼠单克隆抗体(1 ∶1 000)作为内参。相应二抗(1 ∶5 000),室温孵育1 h。应用Odyssey成像系统扫膜,Gel-Pro Analyzer凝胶定量分析软件分析蛋白条带吸光度值。
1.5 F-actin染色 将NG组、HG组、HO组、HG+IQGAP1质粒转染组、HG+空质粒转染组细胞分别培养于多聚赖氨酸涂层的盖玻片上,磷酸缓冲盐溶液冲洗,4%多聚甲醛室温固定15 min,Triton X-100穿透,磷酸缓冲盐溶液冲洗后,异硫氰酸荧光素(fluorescein isothiocyanate,FITC)-鬼笔环肽(5 μg/mL)4℃孵育过夜,4′,6-二脒基-2-苯基吲哚染核,磷酸缓冲盐溶液冲洗后,抗荧光淬灭甘油封片,荧光显微镜观察细胞内F-actin分布。采用F-actin外周环评分(cortical F-actin score,CFS)评价F-actin分布情况:(1)F-actin形成正常应力纤维为0分;(2)F-actin外周环<1/2胞膜边缘为1分;(3)F-actin外周环≥1/2胞膜边缘为2分;(4)F-actin形成完整外周环为3分;每个视野计数100个细胞,取平均值。
1.6 免疫共沉淀分析 取NG组、HG组、HG+IQGAP1质粒转染组、HG+空质粒转染组细胞,按上海碧云天生物技术有限公司免疫沉淀标准操作流程进行实验。裂解细胞收集蛋白后,加入2 μg IQGAP1兔多克隆抗体,4℃过夜,加入20 μL充分重悬的Protein A+G Agarose,4℃缓慢摇动3 h,2 500 r/min离心5 min,收集沉淀,加入40 μL 1×SDS-PAGE电泳上样缓冲液,100℃煮沸5 min,取样品用于SDS-PAGE电泳,以免疫沉淀条带中Cdc42与IQGAP1灰度值的比值评估Cdc42和IQGAP1的结合情况。
1.7 统计学分析 采用SPSS 17.0软件进行统计分析。计量资料以(x±s)表示,组间比较采用单因素方差分析。以P<0.05为差异有统计学意义。
2 结 果
2.1 葡萄糖刺激对足细胞IQGAP1蛋白表达的影响 (1)高糖刺激48 h后的足细胞IQGAP1蛋白表达水平低于高糖刺激0 h后(P<0.05),见图1及表1。(2)应用不同浓度葡萄糖刺激分化成熟的足细胞48 h,30 mmol/L葡萄糖刺激后足细胞的IQGAP1蛋白表达水平低于5 mmol/L葡萄糖刺激后的表达水平(P<0.05),见图2及表2。(3)葡萄糖刺激48 h后,NG组与HO组IQGAP1蛋白的表达情况差异无统计学意义(P>0.05),见图3及表3。
2.2 高糖刺激对足细胞F-actin骨架分布的影响 NG组及HO组足细胞的F-actin聚集成束,呈微丝样结构,延细胞长轴分布,形成应力纤维。刺激48 h后,HG组足细胞的F-actin排列紊乱,F-actin形成核外周环。HG组的CFS高于NG组(P<0.05),HO组与NG组的CFS差异无统计学意义(P>0.05),见图4及表4。
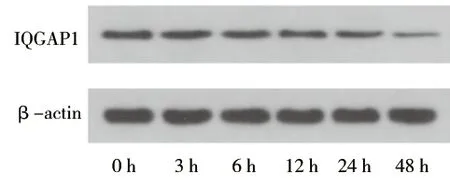
图1 高糖刺激不同时间后足细胞IQGAP1蛋白的表达
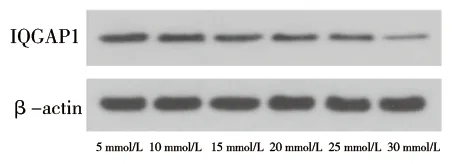
图2 不同浓度葡萄糖刺激48 h后足细胞IQGAP1蛋白的表达
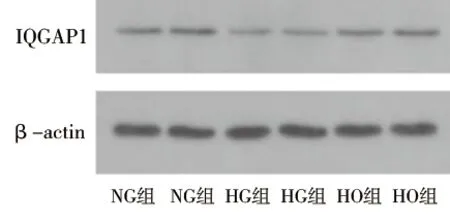
图3 NG组、HG组、HO组IQGAP1蛋白的表达情况

时间nIQGAP1相对表达水平0 h30.56±0.043 h30.55±0.026 h30.53±0.0212 h30.50±0.0124 h30.43±0.0248 h30.23±0.04∗ F值43.843P值<0.001
注:与0 h组比较,*P<0.05。
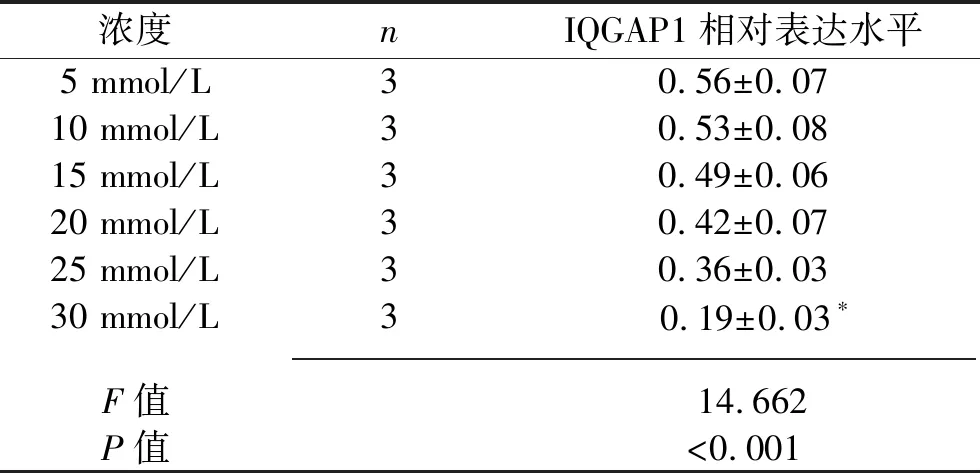
表2 不同浓度葡萄糖刺激48 h后足细胞IQGAP1蛋白的表达情况(x±s)
注:与5 mmol/L组比较,*P<0.05。

表3 NG组、HG组、HO组IQGAP1蛋白的表达情况(x±s)
注:与NG组比较,*P<0.05。
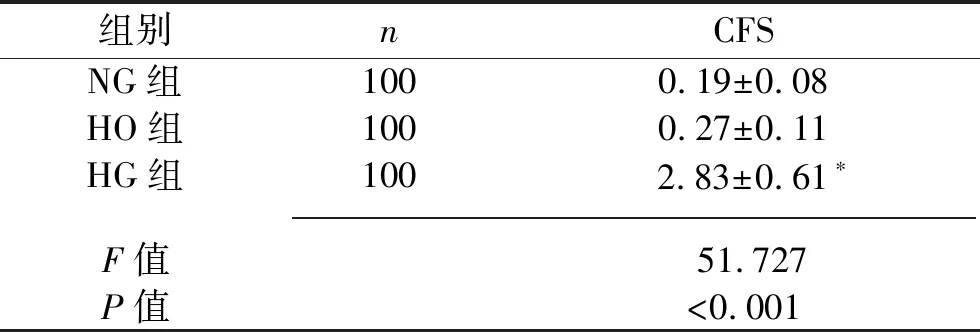
表4 NG组、HG组、HO组足细胞CFS比较(x±s,分)
注:与NG组比较,*P<0.05。
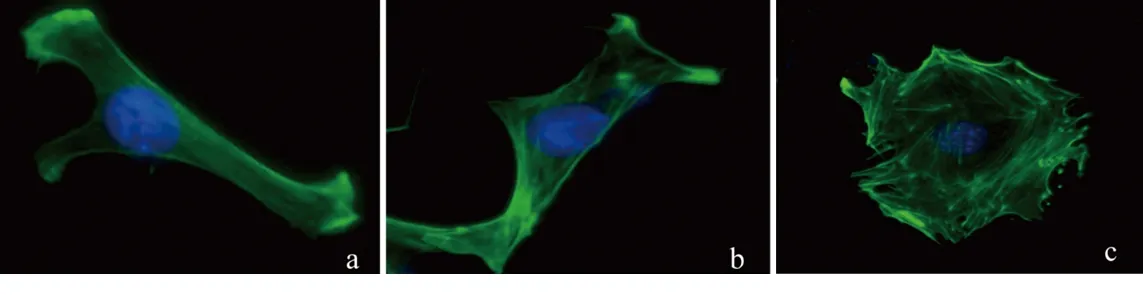
图4 高糖刺激对足细胞F-actin骨架分布的影响(免疫荧光,×400)
2.3 IQGAP1质粒转染对足细胞F-actin骨架分布的影响 FITC-鬼笔环肽染色结果显示,NG组足细胞的F-actin聚集成束,呈微丝样结构,沿细胞长轴分布,形成应力纤维;HG组足细胞的F-actin排列紊乱,F-actin形成核外周环;HG+IQGAP1质粒转染组的足细胞骨架重排部分逆转。HG+IQGAP1质粒转染组的CSF低于HG组(P<0.05),而HG+空质粒转染组与HG组差异无统计学意义(P>0.05)。见图5及表5。
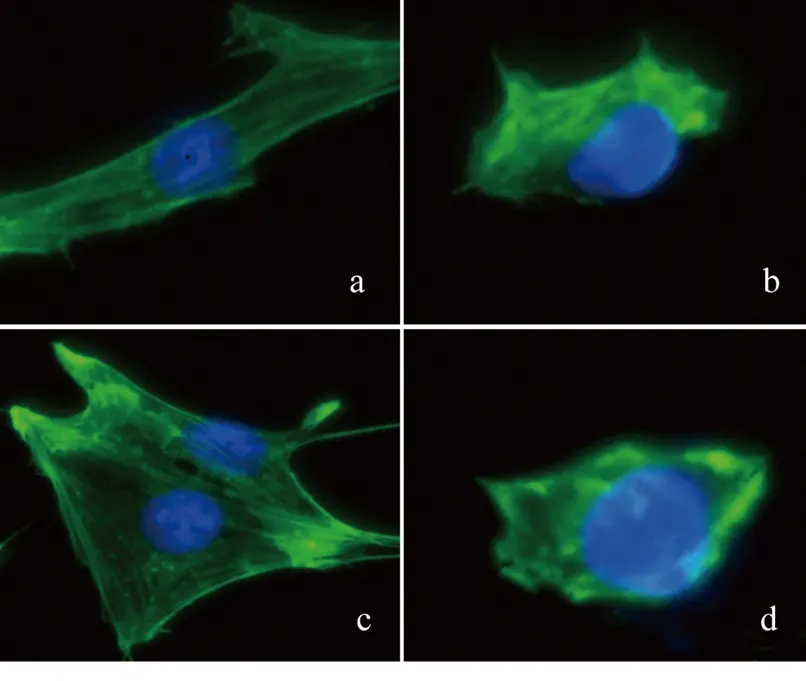
图5 IQGAP1质粒转染对足细胞F-actin骨架分布的影响(免疫荧光,×400)
注:a为NG组;b为HG组;c为HG+IQGAP1质粒转染组;d为HG+空质粒转染组。
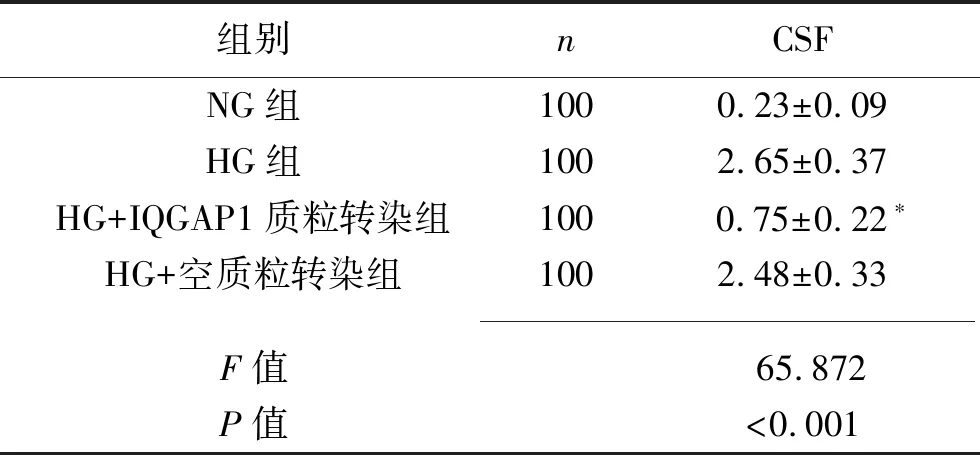
表5 4组转染后足细胞的CSF比较(x±s,分)
注:与HG组比较,*P<0.05。
2.4 IQGAP1质粒转染对足细胞IQGAP1及Cdc42蛋白表达的影响 (1)与NG组比较,NG+IQGAP1质粒转染组的IQGAP1蛋白表达增加(P<0.05),而NG+空质粒转染组的IQGAP1蛋白表达与之差异无统计学意义(P>0.05),提示IQGAP1质粒转染成功。HG组的IQGAP1蛋白表达水平低于NG组(P<0.05),而HG+IQGAP1质粒转染组的IQGAP1蛋白表达水平高于HG组(P<0.05)。见图6及表6。(2)HG组的Cdc42蛋白表达水平低于NG组(P<0.05),而HG+IQGAP1质粒转染组的Cdc42蛋白表达水平高于HG组(P<0.05),HG组与HG+空质粒转染组差异无统计学意义(P>0.05)。见图6及表7。

图6 IQGAP1质粒转染后足细胞IQGAP1和Cdc42蛋白的表达情况
注:1、2、3、4、5、6分别为NG组、NG+IQGAP1质粒转染组、NG+空质粒转染组、HG组、HG+IQGAP1质粒转染组、HG+空质粒转染组。
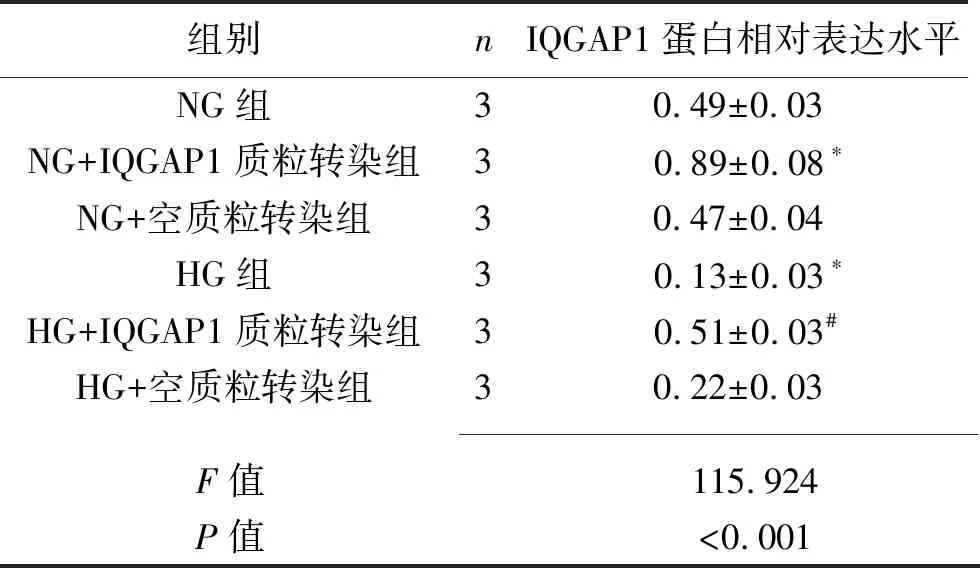
表6 6组足细胞IQGAP1蛋白的表达情况(x±s)
注:与NG组比较,*P<0.05;与HG组比较,#P<0.05。
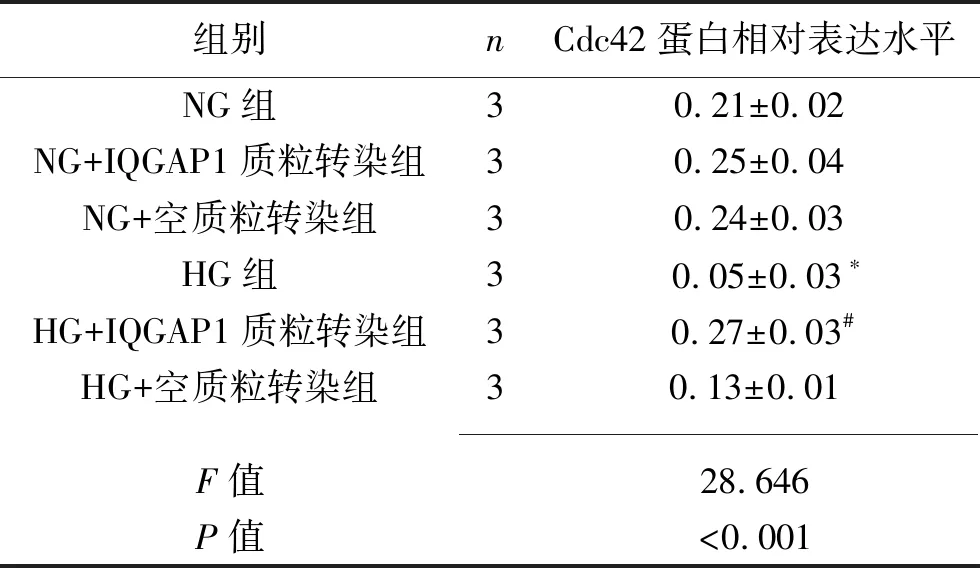
表7 6组足细胞Cdc42蛋白的表达情况(x±s)
注:与NG组比较,*P<0.05;与HG组比较,#P<0.05。
2.5 IQGAP1质粒转染对IQGAP1和Cdc42相互作用的影响 与NG组比较,HG组的IQGAP1与Cdc42结合减少(P<0.05);与HG组比较,HG+IQGAP1质粒转染组的IQGAP1与Cdc42结合增加(P<0.05);而HG+空质粒转染组与HG组的IQGAP1与Cdc42的结合情况差异无统计学意义(P>0.05)。见图7及表8。
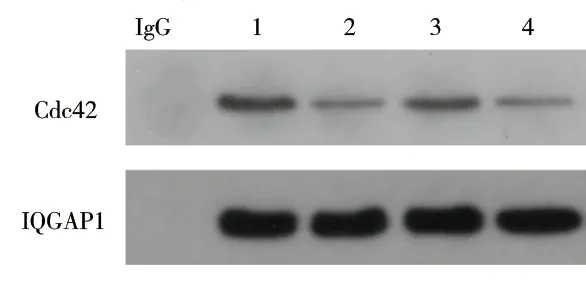
图7 IQGAP1质粒转染后IQGAP1和Cdc42的结合情况
注:1、2、3、4分别为NG组、HG组、HG+IQGAP1质粒转染组、HG+空质粒转染组。

表8 4组足细胞中IQGAP1与Cdc42的结合情况(x±s)
注:与NG组比较,*P<0.05;与HG组比较,#P<0.05。
3 讨 论
足细胞病是一种由多种因素引起,以足细胞的结构和(或)功能损伤为主要特点的肾小球疾病[5],主要表现为足突融合、凋亡或去分化增殖。足突融合是足细胞损伤的特征性微观病理改变,见于多种肾小球疾病,如微小病变型肾病、糖尿病肾病、狼疮性肾炎等[6]。足突融合与足细胞骨架重排密切相关,其中足细胞骨架重排是足突融合的共同通路[2]。然而糖尿病肾病足细胞骨架重排的机制尚不明确。
IQGAP是一种含有多个蛋白结合域的支架蛋白家族,因含有类似RasGTPs催化域系列和4个可与钙调蛋白相互作用的IQ基序而得名。IQGAP1是其中最重要且分布最广泛的成员,含有肌动蛋白结合区、细胞外调节蛋白激酶Ⅱ蛋白结合区、Rho鸟苷三磷酸酶(Rho-guanosine triphosphatase,Rho GTPase)结合区及钙调蛋白结合区[7]。既往研究表明IQGAP1在细胞骨架的调节中发挥重要作用,其可通过调节细胞骨架重排来调节细胞黏附、迁移和极化[8-11]。本课题组的前期研究结果表明IQGAP1参与嘌呤霉素诱导的足细胞骨架重排[4]。但关于IQGAP1在糖尿病肾病领域的研究却很少。本研究结果显示,HG组足细胞的F-actin排列紊乱,F-actin形成核外周环,且其CFS高于NG组(P<0.05),而HO组与NG组的CFS无差异(P>0.05),这提示高糖刺激可引起足细胞骨架重排;高糖(30 mmol/L)刺激后足细胞的IQGAP1蛋白表达水平低于正常浓度(5 mmol/L)葡萄糖刺激后的水平(P<0.05),且高糖刺激48 h后的足细胞IQGAP1蛋白表达水平低于高糖刺激0 h后的水平(P<0.05),提示高糖刺激可下调足细胞的IQGAP1蛋白表达,并在一定程度上呈时间及浓度依赖。此外,HG+IQGAP1质粒转染组的IQGAP1蛋白表达水平高于HG组(P<0.05),提示IQGAP1转染可逆转高糖刺激引起的IQGAP1表达减少;HG+IQGAP1质粒转染组的足细胞骨架重排部分逆转,且其CSF低于HG组(P<0.05),提示上调足细胞IQGAP1蛋白表达后,高糖诱导的足细胞骨架重排改善。以上结果说明IQGAP1蛋白可能参与糖尿病环境中的足细胞骨架重排,我们推测支架蛋白IQGAP1可能通过与某一蛋白结合发挥调节作用。
既往有学者认为,IQGAP1对细胞骨架的调节与Rho GTPase有关,IQGAP1含有可与Rho GTPase家族结合的区域,可通过与Rho GTPase结合调节骨架重排[8-10,12]。Rho GTPase在肌动蛋白细胞骨架调节中发挥着关键作用,目前研究得最多的成员是Cdc42、Rac1、Rho[13]。Cdc42通过诱导肌动蛋白聚合,在足细胞骨架重排中发挥重要作用,研究证实敲除足细胞中Cdc42的编码基因,可抑制肌动蛋白向肾小球足细胞特异性蛋白—nephrin聚集位点集聚,从而导致先天性肾病综合征的发生[14]。有学者对淋巴细胞和神经细胞进行研究发现,IQGAP1、Cdc42、肌钙蛋白可结合形成三联体,发生免疫共沉淀[15-16]。本研究结果显示,HG组的Cdc42蛋白表达水平低于NG组(P<0.05),提示高糖刺激不仅可诱导足细胞IQGAP1蛋白表达减少和足细胞骨架重排,还可下调Cdc42表达。本研究中的免疫共沉淀结果显示,与NG组比较,HG组的IQGAP1与Cdc42结合减少(P<0.05),与HG组比较,HG+IQGAP1质粒转染组的IQGAP1与Cdc42结合增加(P<0.05),提示在高糖条件下小鼠足细胞IQGAP1与Cdc42结合减少,而通过质粒转染增强IQGAP1的表达后,可增加二者的结合,从而改善高糖诱导的足细胞骨架重排。
综上所述,高糖环境可诱导足细胞骨架重排,并减少IQGAP1及Cdc42蛋白表达以及IQGAP1与Cdc42的结合,而IQGAP1可能通过与Cdc42结合而调节细胞骨架重排。IQGAP1/Cdc42结合途径可为糖尿病肾病的防治提供一个新的治疗靶点。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:31:02
大学化学(2021年7期)2021-08-29 12:21:30
通信技术(2019年8期)2019-09-03 08:57:08
生物学通报(2019年3期)2019-06-15 03:35:04
国际呼吸杂志(2019年4期)2019-03-12 01:08:08
中成药(2018年6期)2018-07-11 03:01:04
中成药(2017年8期)2017-11-22 03:18:21
中国学术期刊文摘(2016年2期)2016-02-13 16:01:41
长江蔬菜(2015年3期)2015-03-11 15:10:29
浙江理工大学学报(自然科学版)(2015年7期)2015-03-01 02:54:28
