
银屑病性关节炎的致病基因筛选及生物信息学分析
2019-05-21 03:26何剑戈夏群吴凯楠杨凯锐1贾栋1李明昊
山东医药 2019年8期
何剑戈,夏群,吴凯楠,杨凯锐1,,贾栋1,,李明昊
(1中国人民武装警察部队后勤学院,天津300000;2中国人民武装警察部队特色医学中心;3中国人民解放军92126部队;4天津医科大学;5天津中医药大学)
银屑病性关节炎(PsA)是一种慢性、免疫介导性的炎症性关节病,表现为关节和血管的炎症,包括轴向骨骼的炎症,并与心血管疾病的病死率增加有关[1]。银屑病关节炎与不同程度的残疾有关,延误诊断还可增加死亡风险,因此,早诊断、早治疗可预防关节破坏及改善预后[2,3]。目前还没有公认的血清学筛查方法用于该病的早期诊断。PsA病因尚不清楚,可能是遗传、免疫和环境因素相互作用的结果。PsA有明显的遗传因素,许多基因都与该病的易感性有关[4]。基因芯片技术有助于确定某种疾病特异的诊断标志物或关键的致病基因[5]。生物信息学是分析处理生物分子信息、揭示生物分子信息内涵的一种技术[6],可在生物医学领域获取大量信息,用于遗传性疾病的预防、诊断和治疗相关研究[7]。2018年5~10月,本研究以PsA基因芯片数据作为研究对象,筛选PsA的致病基因并进行生物信息学分析。现报告如下。
1 材料与方法
1.1 主要实验材料 本研究所用芯片数据来源于美国生物信息技术中心(NCBI)的公共基因芯片数据(GEO),芯片数据编号为GSE61281,基于GPL6480平台(Agilent-014850 Whole Human Genome Microarray 4x44K G4112F),疾病类型为PsA和银屑病[8]。该芯片数据有52个样本,包括12个来自正常人的外周血样本(对照组)、20个PsA患者的外周血样本(试验组)和20例银屑病患者的外周血样本(试验组)。本研究分析12个对照组外周血样本和20个PsA患者外周血样本的芯片数据。
1.2 差异表达基因的筛选及优化 利用R编程软件对系列矩阵文件进行标准化处理[9]。将芯片探针名转化为基因名。对于多个探针与同一基因对应者,基因表达量取多个探针上的均值。预处理前芯片数据共有41 092个探针符号,通过对原始芯片数据进行标准化、删去无相应基因符号探针及归一化处理,得到19 595个没有重复的基因符号。用R软件limma工具包筛选对照组与PsA试验组芯片数据的差异表达基因[10],选择校正P<0.05且差异表达倍数为1.5倍以上者作为差异表达基因。在GLAD4U数据库(该数据库通过检索PUBMED文献,可明确基于现阶段科学认知水平较可信的致病基因,即已知基因[11])中输入关键词“psoriatic arthritis”来检索PsA的已知基因。但仍有很多与PsA致病相关且未研究过的基因,在GLAD4U数据库中无法检索到。为排除在样本收集、芯片测序过程中发生的污染及在芯片数据分析时发生的偏差,需要对初筛的差异基因进行优化提升。将GLAD4U数据库中得到的已知基因作为训练集,将通过R软件筛选得到的差异表达基因作为测试集,通过ToppGene数据库(该数据库根据两组基因在功能上的相似性对疾病候选基因进行优化和排序)优化测试集,最终选择综合P<0.01、综合得分>0.6的基因作为差异表达基因中与PsA相关性更高的基因,将这些基因和训练集内的已知基因共同纳入致病基因。
1.3 致病基因的富集分析 利用David(https://david.ncifcrf.gov)在线工具对致病基因进行GO富集分析,选择经过Benjamini校正后P<0.05的基因本体[12]。利用Kobas数据库对致病基因进行KEGG分析[13],选择经过Benjamini校正后P<0.05的通路作为致病基因的主要富集通路。
1.4 致病基因PPI网络的构建 利用String数据库构建PsA致病基因的PPI网络,并建立可视化PPI网络图,计算PPI网络中各个基因的节点度(某致病基因与其他致病基因有直接联系的数目),可获得在PPI网络中与其他致病基因有密切联系的基因。
2 结果
2.1 差异表达基因的筛选及优化结果 利用R编程语言的limma工具包筛选差异基因,共获得差异表达基因104个,其中表达上调基因83个、表达下调基因21个。GLAD4U数据库中检索获得已知基因59个。将已知基因作为训练集,差异表达基因作为测试集,通过ToppGene数据库优化测试集,筛选出15个与已知基因功能相似的基因。将这15个基因与已知基因共同纳入PsA的致病基因。差异表达基因中与PsA高度相关(得分较高且差异显著)的基因为CXCL10、LYN、JAK1、CARD11、ANXA1。
2.2 致病基因的富集分析结果 经过David在线工具进行GO富集分析,得到199个致病基因富集的基因本体。P<0.05、基因富集个数排列前15个的基因本体见表1。经过KOBAS在线工具进行KEGG富集分析,得到94个致病基因富集的KEGG通路。P<0.05、基因富集个数排列前15个的KEGG通路见表2。
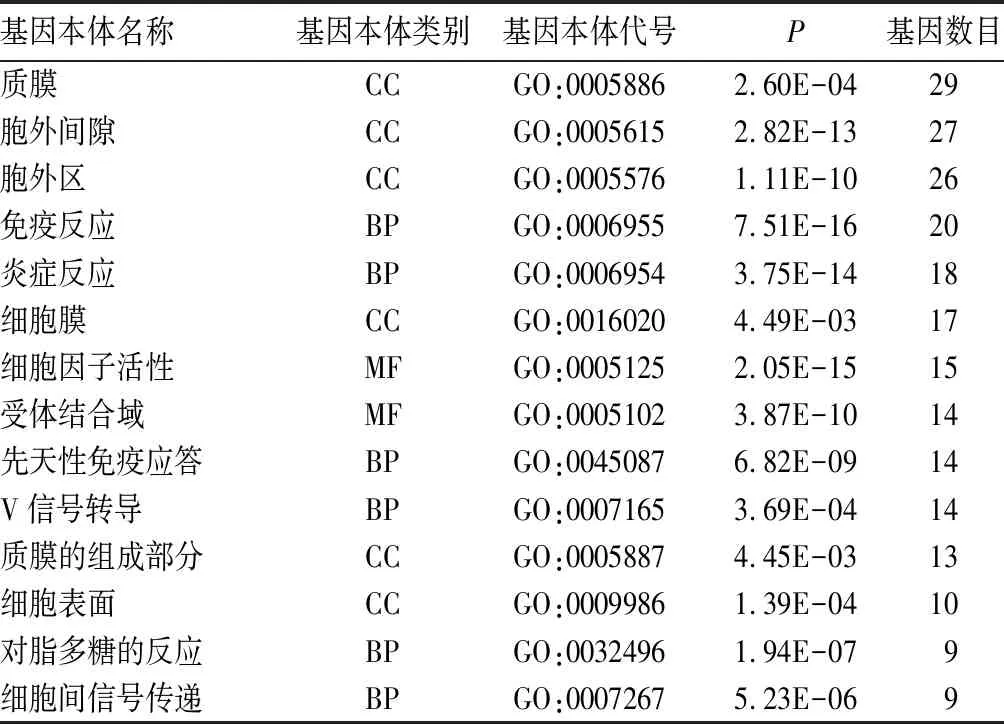
表1 致病基因的GO富集分析结果
注:MF为分子功能;BP为生物过程;CC为细胞组成。
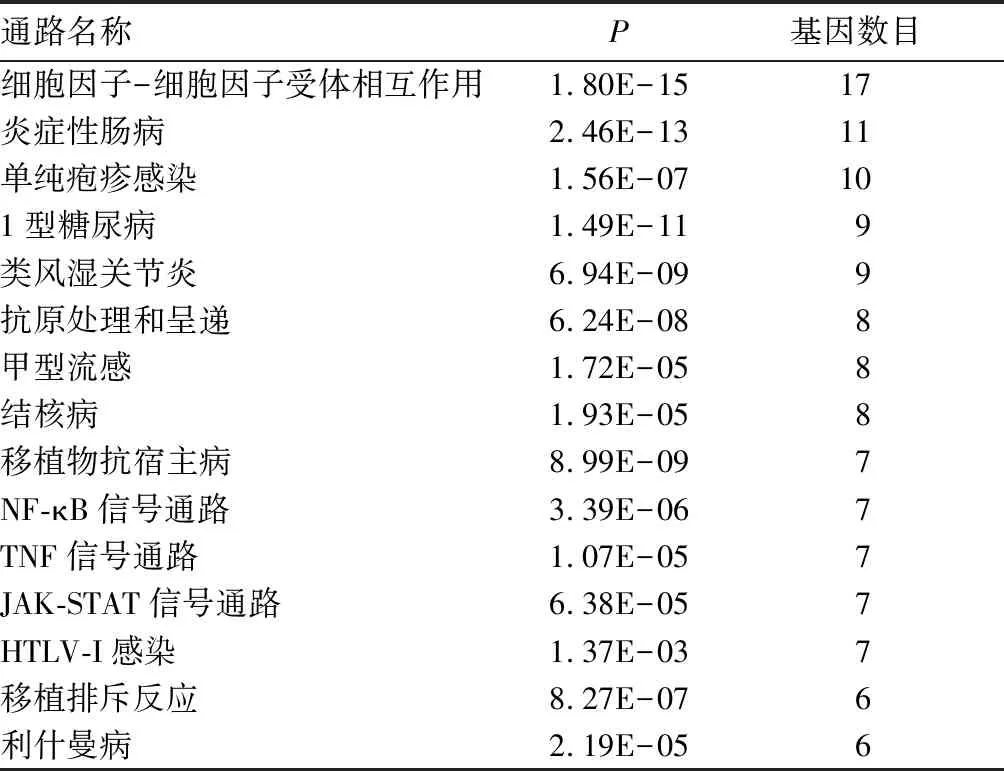
表2 致病基因的KEGG通路富集分析结果
2.3 致病基因的PPI网络 致病基因的PPI网络中TNF节点度较大,即TNF在PPI网络中与其他基因的直接作用较多,联系密切。IL1B、IL13、IL17A、CCL2等基因也有较大的节点度。见图1。
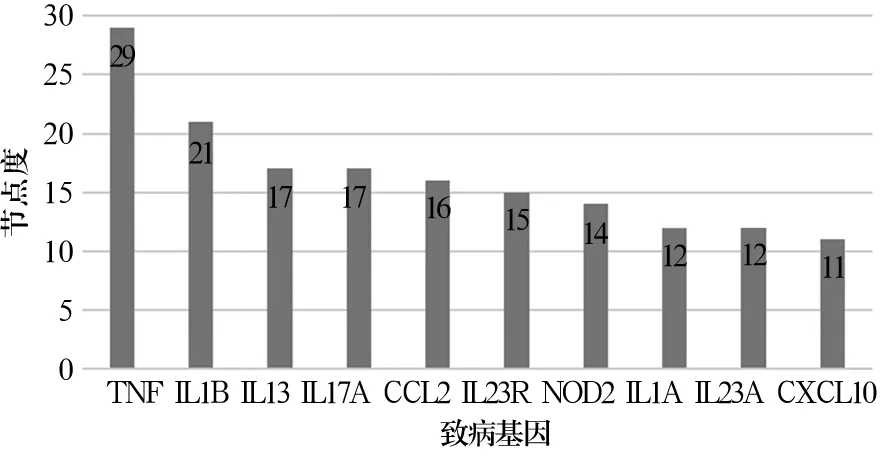
图1 PPI网络中节点数排名前10位的致病基因
3 讨论
通过对来自GEO数据库基因芯片的分析,可以了解某些基因在疾病发生过程中的作用机制。然而,基因芯片分析得到的差异表达基因对进一步探究发病机制的指导作用并不明确。本研究从GLAD4U数据库中检索出与PsA发病密切相关的已知基因,并将其作为训练集,将分析基因芯片得到的差异基因作为测试集,通过ToppGene数据库优化测试集,最终选择综合P<0.01、综合得分>0.6的基因作为差异表达基因中与PsA相关性更高的基因,将这些基因和训练集内的已知基因共同组成致病基因。差异表达基因中与PsA高度相关的基因为CXCL10、LYN、JAK1、CARD11、ANXA1。
JAK-1是优化的差异基因中得分较高(0.920 833 852)且显著(P=4.01E-05)的基因。JAK家族(JAK-1、JAK-2、JAK-3、TYK-2)在免疫调节细胞因子的细胞内信号转导中发挥重要作用。JAK-3与JAK-1结合对IL的信号转导起重要作用。IL广泛参与淋巴细胞活化、功能和增殖。JAK-3主要表达于造血细胞,包括NK细胞、胸腺细胞和活化的T、B淋巴细胞。JAK-1可与其他JAK家族成员结合,发出其他细胞因子的信号。因此,抑制JAK蛋白是调节T淋巴细胞、B淋巴细胞、巨噬细胞和NK细胞免疫应答有力的靶点[14]。PsA与炎症和免疫反应相关,JAK-1的异常表达有可能与PsA的发生相关。
CXCL10基因在优化的差异基因中得分排名第二。CXCL10蛋白可与受体CXCR3结合,通过激活和招募白细胞调节免疫反应。最近研究显示,CXCL10在多种自身免疫性疾病(如风湿性关节炎、系统性红斑狼疮、干燥综合征、系统性硬化症和特发性炎性肌病等)患者的血清和(或)组织中表达有所增加。本课题组最近研究表明,CXCL10在骨破坏中也发挥一定作用。除了趋化效应外,CXCL10还可能具有多效性功能。进一步研究这种趋化因子的功能及CXCL10与其他细胞因子、趋化因子之间的相互作用,有助于为多种自身免疫性疾病探索治疗靶点[15]。在本研究中CXCL10基因综合得分较高,提示其与PsA的发病高度相关。有研究表明CXCL10可能涉及到PsA的发病机制,认为它是预测银屑病的一种生物标志物[16]。本研究结果与之一致。
本研究对致病基因进行富集分析,结果发现在炎症、免疫和信号转导等相关的生物过程中富集的基因较多且较显著,如免疫反应、炎症反应、固有免疫反应、信号转导、对脂多糖的应答等。有学者[17,18]曾报道PsA是一种炎症和免疫性疾病并且与遗传易感性、环境因素有关。所以炎症反应和免疫功能异常可能是PsA发病的重要病理机制。KEGG分析结果发现,PsA致病基因主要富集的通路是细胞因子与细胞因子受体相互作用、炎症性肠病等。在细胞因子与细胞因子受体相互作用通路中,富集较多致病基因的通路为IL6/12样细胞因子通路、IL17样细胞因子通路、TNF家族通路。
血清TNF-α水平与PsA病情活动CASPAR评分和各项炎症指标密切相关,是PsA发病的重要机制[19]。IL-17A在炎症相关的关节炎和(或)皮肤疾病的发病中起关键作用。在明显的关节炎症之前,IL-17A可通过直接激活破骨细胞前体来诱导病理性骨吸收[20]。在对致病基因的PPI网络分析中,我们发现TNF、IL1B、IL13、IL17A、CCL2等基因也有较大的节点度。进一步说明这些信号通路中富集的基因可能成为PsA诊断和治疗的潜在靶点。
本研究通过对PsA的基因芯片进行生物信息学分析,找到了与PsA致病相关的核心基因及重要通路,为PsA的发病机制研究提供了重要的理论和数据支撑。对这些核心基因及通路进一步研究有助于深入揭示PsA的发病机制。
猜你喜欢
环球时报(2022-09-29)2022-09-29
现代临床医学(2021年4期)2021-07-31
发明与创新(2021年17期)2021-07-05
今日农业(2021年4期)2021-06-09
军民两用技术与产品(2021年12期)2021-03-09
知识文库(2020年5期)2020-05-11
中西医结合心脑血管病杂志(2016年20期)2016-03-01
中国实用医药(2016年1期)2016-01-11
医学研究杂志(2015年12期)2015-06-10
中国药房(2010年12期)2010-02-12
