
水通道蛋白5在干燥综合征中的作用综述*
2019-05-14 01:56沙明慧吴素玲南京中医药大学南京009南京市第一医院南京000
药学与临床研究 2019年2期
沙明慧,吴素玲**南京中医药大学,南京 009;南京市第一医院,南京 000
水通道蛋白5(AQP5)是一种膜转运蛋白,最先是从大鼠颌下腺的cDNA中扩增出来的[1],1996年被Lee等成功克隆,近年来关于其相关信号通路研究备受关注。干燥综合征(SS)是一种以侵犯外分泌腺为主的慢性炎症性自身免疫性疾病,临床多以口眼干涩为主要表现,可累及肺脾肝肾、甲状腺等多脏器,好发于40~50岁女性,男女比例为1∶9。目前病因及发病机制并不十分明确,现代医学尚无理想的治疗方案,主要是对症缓解干燥症状,阻止、延缓病情进展,治愈率较低,可严重影响患者健康水平及生活质量。SS是现代医学的一大疑难病问题,调控AQP5有可能是改善SS症状的关键点。
1 AQP5的生物学特征
1.1 AQP5的结构
水通道蛋白家族(aquaporins,AQPs,结构见图 1)是动植物细胞膜上的水孔蛋白簇,目前已发现有13种水通道蛋白(AQP 0-AQP 12),是一种整体膜蛋白。其中AQP5由六螺旋多肽链组成,可6次跨膜,分子量约28 kD,包含有5个襻,细胞外为亲水性A、C、E环,细胞内为疏水性B、D环。其中A、C、E环的氨基端和羧基端位于细胞内,两者重复部分构成AQP,分子前后蛋白基序相似,形成了以整个AQP分子为中心的对称镜像结构。而B环和E环均具有NPA基本序列(Asp-Pro-AIa,天冬酰氨酸-脯氨酸-丙氨酸基序),B、E 环在NPA处向细胞膜内凸起、褶皱形成一个单孔凹穴,使AQP5呈“沙漏样”结构、钩嵌在细胞膜上,4个单体形成四聚体以凹穴为中心排列,构成了腺体的特异性分泌通道,有高渗性特征,在水转运功能上起到了决定性作用。凹穴内侧的A-73和C-189残基若缺失或被替代,均可导致水液转运障碍。此外,AQP5是汞剂敏感性通道,182位的半胱氨酸可与汞剂结合、抑制水通道功能。AQPs中几乎所有亚型及同源蛋白中都具有NPA这个典型氨基酸序列。人的AQP5氨基酸序列与大鼠、小鼠的同源性分别为98%、90%,与大鼠基本无差异性[2]。
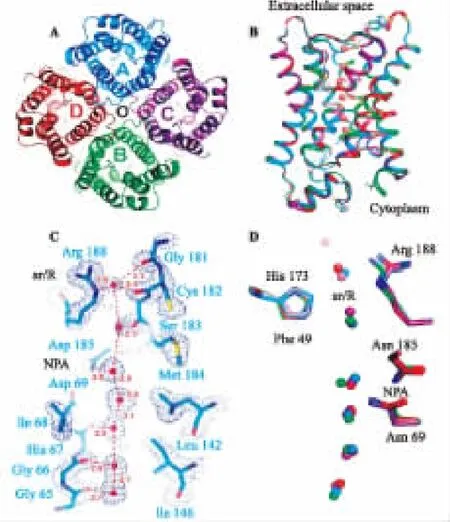
图1 AQP5的晶体结构[3]
1.2 AQP5的分布
AQPs因表达组织及具体部位的不同,有组织和细胞特异性,各自发挥不同的生理特征。其中AQP5主要分布于泪腺、涎腺以及角膜、上呼吸道、肺泡Ⅰ型细胞等各类分泌性上皮细胞及腺体处。在某些组织器官中AQP5也有高表达,如肺、肾;在脑、胰腺、小肠等器官及组织中未见表达。此外,在正常情况下,AQP5存在于细胞囊泡中,受到体内外某些刺激后出现由囊泡向细胞顶膜 “转位”并发生融合的现象,即AQP5的重新分布特性。李磊磊等[4]表明,AQP5可能参与了肺胀阳虚水泛证水液代谢失衡的过程。任鸿昌[5]实验结果表明,胰蛋白酶导致的急性肺损伤后期,AQP5在肺组织中表达逐渐下降,促进了肺水肿形成,肺损伤程度相应加重。秦崧[6]发现,AQP5高表达于肺、呼吸道黏膜下腺腺体细胞腔,为适应极干旱荒漠环境提供了蛋白水平依据。在气道高反应过程中,AQP5的表达下调,粘蛋白MUC5B高表达,气道粘液高分泌。Ma T等[7]发现,AQP5基因敲除小鼠较野生型小鼠肺内渗透压依赖的水转运下降90%。总之,AQP5的异常分布、转运对水液代谢、维持渗透压平衡和腺体分泌等有重大影响。
2 AQP5的生物学功能
2.1 AQP5及其相关信号通路
2.1.1 TLR4/MyD88/NF-κB/AQP5信号通路 Toll样受体4(Toll like receptor,TLR4)可通过与核因子 NF-κB(nuclear factor of kappa B)相应配体结合,激活NF-κB和丝裂原活化蛋白激酶信号通路,来促进各类炎症因子的释放。NF-κB的激活可以使 AQP5低表达[8,9]。而髓样分化因子 88(myeloid differentiation factor 88,MyD88)是TLR信号转导途径中主要的接头蛋白,故可调节TLR4/NF-κB信号通路。
2.1.2 MBD2/AQP5信号通路 去甲基化酶MBD2(methyl-CpG binding domain2)是一种特异性结构域蛋白,甲基化的胞嘧啶-磷酸-鸟嘌呤(CpG)是MBD2的启动子,MBD2与其结合受到启动后,具有去甲基化功能,可使DNA甲基化,从而参与AQP5基因表达受抑,具体作用及调控机理尚不清楚。故MBD2/AQP5信号通路可能是AQP5表达的重要调控机制之一。
2.1.3TNF-α/TNFR/IKK/NF-κB、MKK/p38/c-Fos、TRAF6/JNK/c-Jun信号通路 肿瘤坏死因子TNF-α通过与其受体1(TNFR1) 相结合形成的复合物可激活 NF-κB;IκB 激酶(IKK)的激活可使 IκB磷酸化、降解,促使 NF-κB激活,从而抑制AQP5蛋白和mRNA的表达。p38、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)与 c-junN-末端激酶(JNK)并称为应激活化的蛋白激酶(SAPKs)。NF-κB 是细胞外调节蛋白激酶 (extracellular regulated protein kinases,ERK)/MAPK信号通路的下游调节因子。JNK属于MAPK家族的一种信号转导蛋白,受其上游激酶激活后,可传导ERK、JNK、p38等通路,调控细胞迁移。p38蛋白与JNK相似,同属MAPK家族亚类,p38-MAPK被上游激活后可磷酸化IκB,并将之降解,IκB、NF-κB 解聚,NF-κB 被激活,产生促炎效应,可调节细胞增殖、分化。NF-κB、JNK都是由TNF-α信号通路介导的。高表达的ERK、p38和JNK均抑制AQP5表达[10]。
以上各种信号通路均可调控AQP5,使AQP5转录抑制,故在功能上有相似之处,但属于不同途径来调节水液代谢和腺体分泌。目前对于影响AQP5的信号转导通路的研究及认识尚且不足,但为AQP5与SS之间的研究提供了思路,其主要区别为基因水平与分子水平的不同研究方向。
2.2 AQP5的调节机制
AQP5是一种利用渗透压差有效促进水分子快速转运的跨膜转运蛋白,在生物体内有长时调节和短时调节两种调节机制。长时调节主要是指通过某些方法干预可促使AQP5 mRNA和蛋白水平的上升,以及提升其跨膜转移的生物学功能,即调节AQPs含量作用。例如,Smith JK[11]将IFN-α加入人的唾液上皮细胞,经过1 h刺激,可显著上调AQP蛋白和mRNA。短时调节则是指在内环境等因素调节下,可以改变AQP5的生物学特征和功能,即调节AQPs活性作用,目前公认磷酸化机制和穿梭机制参与了AQP5的短时调节。AQP5结构的主要作用是蛋白激酶 A(protein kinaseA,PKA)和蛋白激酶 C(protein kinase C,PKC)介导的磷酸化,AQP5 含有PKA和PKC磷酸化的同源序列,使AQP5结构改变,膜通透性增加,从而产生“快速闸门”效应,证实了AQP5在水液的转运、门控等方面发挥着重要的调节作用,而AQP5的磷酸化功能是依赖cAMP实现的,可以通过调整cAMP活性来节制磷酸化作用。穿梭机制则是指通过改变内环境中Ca2+浓度、细胞结构等条件,在某些因素影响下,可使正常存在于细胞内囊泡中的AQP5转位,从而改变细胞膜通透性及功能性水通道数目,表明AQP5具有调节水分子重新分布的生物学功能。例如,在SS相关的AQP5表达下调研究中,用乙酰胆碱刺激腺体腺泡细胞表面M3受体,并与其相结合来启动二级信号传导效应,促使Ca2+浓度增加,诱导胞质内的AQP5经囊泡向腺泡细胞顶膜转运。上述现象主要通过增强电解质活跃转运及诱导AQP5“转位”等途径实现的,都可以增强腺泡细胞的水分子输出,从而共同参与AQP5的水跨膜转运调节功能。
2.3 AQP5与其他疾病相关性
AQP5的缺失及功能失调会引起多系统疾患。在角膜疾患中,AQP5功能失调会导致角膜水肿、干眼症。而Ⅰ型糖尿病引起的口腔干燥症、老年性口干症、头面部放疗后口干症,均与SS发病机制及临床表现相似。AQP5在类风湿关节炎(Rheumatoid arthritis,RA)小鼠中明显表达上调,并与 CD90、CD44在小鼠骨髓间充质干细胞膜上共表达[12],这说明AQP5与RA的发病机制也有一定关系。在呼吸系统、温燥条件下和肺炎支原体(mycoplasma pneumoniae,MP)感染后均能引起肺部AQP5低表达[13];在慢性阻塞性肺疾病大鼠模型中,AQP5在气道中的表达明显减少(P<0.01)[14]。刘春枝等[15]研究发现,新生儿呼吸窘迫综合征(respiratory distress syndrome of newborn,RDSN)患者的AQP5水平明显上调,且与病情严重程度、胎龄及预后呈一定相关性。中医药方面发现,参七化痰方、三仁汤、竹沥等均可通过调节AQP5表达水平,阻碍粘液高分泌,从而改善肺通气状态[14,16];此外,AQP5与气道高反应相关,故与哮喘发病等也有一定关系。AQP5在皮肤系统主要表达于皮肤颗粒层角质细胞的细胞膜,对维持皮肤细胞间水稳态发挥重要影响。目前多项研究显示,AQP5失调与梅尼埃病、高血压病、糖尿病肾病、变异性鼻炎等的发病机制也具有一定相关性[17-19]。也有学者认为,AQP5可能还参与了近排卵期卵巢囊内液体动态平衡。另有研究显示,AQP5在乳腺癌、胃癌、肝癌、肺癌、宫颈癌、结直肠癌等人类多种肿瘤中过表达,由此证明AQP5的主要功能有调节肿瘤细胞的迁移、增殖和凋亡,与淋巴转移及预后不良也具有一定相关性[20-22]。
3 AQP5在SS发病中的作用
3.1干燥综合征概况
干燥综合征是一种主要侵犯泪腺、唾液腺等外分泌腺体的炎症性自身免疫性疾病,具有高度淋巴细胞浸润,腺体细胞受损的特征。属中医“燥证”“燥痹”范畴。中医认为,燥痹虽以阴虚津亏为本,然而津液输布障碍亦是关键。调控AQP5正是符合中医疏通布津通道之法,旨在促进津液输布通达,得以濡养四肢百骸及皮肤、官窍。临床研究已发现,SS患者泪液中AQP5的表达量较健康人群明显升高,目前认为是慢性淋巴细胞浸润和破坏导致泪腺腺泡细胞受损、AQP5漏入泪液中导致。同时AQP5表达量的减少,可引起唾液流量减少、粘稠度增加,导致分泌异常。结合AQP5的结构功能特性,不难得出AQP5与SS发病具有高度相关性。
西医治疗方面,干燥综合征目前尚无根治方法。一方面主要是人工唾液、泪液的替代治疗,可对症缓解口眼干燥症状,或者用毒蕈碱胆碱能受体(muscarinic receptors)激动剂,刺激外分泌腺分泌;另一方面,SS患者应及时给予系统治疗,包括以下几点:①免疫抑制和免疫调节治疗:羟氯喹(hydroxychloroquine,HCQ)200~400mg·d-1[6~7mg·(kg·d)-1]常作为初始治疗,其他有甲氨蝶呤(methotrexate,MTX)7.5~15 mg·w-1[2~3mg·(kg·w)-1],环磷酰胺(CTX)50~150 mg·d-1或 0.5~1 g·(m2·4w)-1,来氟米特(leflunomid,LEF)10~20 mg·d-1,硫唑嘌呤(azathioprine,AZA)50~100 mg·d-1[1~2 mg·(kg·d)-1],环孢素 A(cyclosporinA,CsA)2.5~5 mg·(kg·d)-1,吗替麦考酚酯(mycophenolate mofetil,MMF)0.75~2 g·d-1。 ②糖皮质激素治疗:根据病情决定用量,泼尼松10~60 mg·d-1,必要时甚至需要糖皮质激素冲击治疗。③生物制剂:目前有抗CD20单克隆抗体(利妥昔单抗,rituximab)、抗CD22单克隆抗体(依帕珠单抗,epratuzumab)等治疗活动性SS有一定临床疗效。④其他:非甾体抗炎药(NSAIDs),如洛索洛芬钠片、双氯芬酸、塞来昔布和美洛昔康等。
3.2AQP5与SS的关系
有关学说认为,AQP5的亚细胞分布及转运异常可能是SS发病的的重要机制之一。在炎症浸润的NOD小鼠颌下腺腺泡中,AQP5在其顶质膜表达减少而在基底膜表达增加。Konttinen YT等[23]发现,AQP5在SS模型小鼠的涎腺上皮表达减少或消失,在肌上皮表达增强。此后在pSS患者中也验证了上述结果。且目前已有临床试验[24]发现,在抗肿瘤坏死因子-α(TNF-α)抗体(infliximab)治疗 pSS 前后,AQP5 的分布由腺泡细胞表面的腔面、基底侧面集中转移至腔面,并据此推测恢复AQP5在腔面的适当分布,可以增加无刺激状态下SS患者的唾液流率。通常认为,AQP5主要是在涎腺腺泡细胞的顶质膜上定位,而目前已发现,在SS患者及小鼠模型的涎腺中,AQP5在顶质膜上分布减少,而在基底膜上增加,借此AQP5调节水分子重新分布功能发挥作用,这说明AQP5在SS的水转运及腺体分泌过程中,至少在刺激唾液分泌中有重要生理特性。而自身IgG抗体及其介导的抑制作用、体内渗透压、cAMP、PKA等因素都已被证实参与了AQP5以上调控过程。总之,AQP5的缺失、异常分布或转运对SS患者的水液代谢和腺体分泌过程都有重大影响。另外,免疫斑点实验证明,AQP5在分泌粘液的颌下腺及舌下腺的表达远较单纯分泌浆液的腮腺丰富。据此又推测,AQP5从本质上有粘液水合作用及溶解作用,而不是液体的分泌作用,但此观点仍需进一步研究证实。
目前众多研究已发现,AQP5及其相关信号通路在SS中可能发挥重要作用。从动物实验中发现,在SS动物模型中NF-κB表达增加,进而激活炎症反应,引起血清中相关炎症因子 TNF-α、IL-6、IL-10、IL-1β 和 IFN-γ 等的增加,可加剧SS小鼠腺体细胞的损害[25,26]。而 NF-κB是 AQP5的上游启动序列之一,在原发性干燥综合征 (Primary Sjogren's syndrome,pSS)的患者中,唇腺、涎腺细胞中的 TLR4和 MyD88较正常人群呈高表达状态。浸润性单核细胞、腺泡细胞和导管上皮细胞中也同样发现了TLR4和MyD88的表达。故不难得出结论:TLR4/MyD88/NF-κB/AQP5信号通路与 AQP5高度相关,并与SS的发病具有一定相关性。近年来亦有研究表明,AQP5及其相关信号通路对颌下腺炎症具有明显调节作用。据临床观察,放疗后患者唾液腺损伤导致的口干症与SS临床表现大致相仿,而AQP5的低表达与SS发病密切相关。李莹[27]已证实了因γ射线辐射引起的AQP5表达降低,从而致使SS的分子机制:在电离辐射致伤人颌下腺(human submandibular gland,HSG)的细胞中,MBD2可以促进AQP5的表达。AQP5的结构特征决定了一旦AQP5表达量减少、缺失及分布异常都可使水转运失代偿,腺体分泌减少,从而出现口干眼干等症状。故MBD2/AQP5信号通路可能在SS患者颌下腺的水分子跨膜转运及腺体分泌过程中发挥重要作用。赵红玉等[10]在蛋白印迹实验中检测到SS模型小鼠颌下腺细胞中NF-κB、p38、c-Jun三种蛋白明显高表达,同时AQP5低表达, 而且三者分别是 TNF-α/TNFR/IKK/NF-κB、MKK/p38/c-Fos、TRAF6/JNK/c-Jun信号通路发挥作用的关键蛋白,故提示以上三条信号通路可能通过抑制AQP5表达参与了SS的发病机制。同时该作者证实,增液汤对这三种关键蛋白均有抑制作用,进一步提示增液汤可能通过调控这三条信号转导通路,上调AQP5的表达,从而抑制炎症及免疫反应,达到治疗干燥综合征的目的。研究也发现,IL-1、IL-6、TNF-α、脂多糖(LPS)等都可激活JNK通路;另外,毒蕈碱M受体、肾上腺素受体介导的作用途径等也均可促进AQP5表达,改善SS患者唾液分泌[28]。故不难得出结论:通过调控AQP5及其相关信号通路来治疗SS,是目前及未来的重点研究方向。
3.3 AQP5与SS的相关中医药研究现状
既往动物实验显示,麦冬多糖可有效降低颌下腺淋巴细胞浸润度,上调颌下腺AQP5和VIP mRNA的表达,即麦冬多糖对NOD小鼠颌下腺具有保护作用[29]。复方制剂津血源颗粒长期研究的结果提示,其可明显调高SS模型鼠唾液腺细胞膜AQP5蛋白的表达,且使其分布均匀[30];进一步研究后认为,津血源增加唾液分泌(养阴生津作用)的机制可能是通过毒蕈碱M受体(尤其是M3受体)、肾上腺素受体介导的作用途径,增加唾液腺细胞膜AQP5的表达,从而促进唾液分泌。中医清燥布津法有明显抑制腺体破坏、改善唾液分泌功能的作用,其作用机制可能与抑制免疫炎症、促进AQP-5在小鼠颁下腺的表达有关[31]。李碧霞[32]发现,利用乌梅喷雾剂干预2周后,能使大鼠颌下腺AQP5基因明显过表达,促进水跨膜运动。铁皮石斛可通过调节SS患者TNF-α、IL-1β和AQP5mRNA的表达,从而改善腺体的淋巴浸润状态,因而对SS具有治疗作用[33]。白芍总苷可通过上调AQP5及其mRNA的表达,改善NOD小鼠颌下腺的病理损害[34]。甘皂颗粒可通过上调AQP5的表达,降低细胞内Ca2+离子的浓度,抑制SS模型大鼠过度增强的免疫反应,减少颌下腺中淋巴细胞浸润,缓解炎症因子浸润,改善唾液腺分泌功能,从而缓解SS口腔干燥的症状[35]。李晓萌等[36]实验,利用AQP5荧光素酶基因启动子筛选到的人参皂苷Rb1药物,证明对AQP5基因有上调作用,并利用动物模型证实Rb1通过促进SS鼠唾液腺的分泌,减轻颌下腺炎症,上调颌下腺AQP5的蛋白表达,从而改善口腔干燥症状。结合动物实验[37]结果,PCR显示补骨脂盐炙后与补骨脂生品比较,AQP5mRNA表达明显升高(P<0.05),提示补骨脂盐炙后药物燥性有所缓解,可能与其对体内AQP的基因表达调控有关。
综上所述,AQP5在SS炎症发展、免疫调节、水分子转运及腺体分泌中发挥重要作用,调控AQP5可能是干燥综合征药物治疗作用的重要靶点,而上述中医药治疗SS已有成效,未来有很大研究开发空间,尚需临床及实验开发相关阻滞剂或基因替代疗法,这可能为许多与炎症、体液代谢有关的疾病提供治疗的新途径。而关于AQP5其上下游调控机制尚待进一步探讨和阐明。
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07
波谱学杂志(2021年3期)2021-09-07
汽车维修与保养(2021年8期)2021-02-16
实用口腔医学杂志(2020年5期)2020-12-07
儿童故事画报·智力大王(2020年3期)2020-04-26
中国医科大学学报(2020年4期)2020-01-08
中国医学影像技术(2019年7期)2019-08-01
作物研究(2017年2期)2017-06-01
浙江医学(2015年2期)2015-01-18
汽车与新动力(2013年1期)2013-03-11
