
干旱胁迫对樟树幼苗光合特性和水分利用的影响
2019-05-13 12:58罗彬莹刘卫东吴际友魏志恒陈明皋廖德志黄小飞
中南林业科技大学学报 2019年5期
罗彬莹,刘卫东,吴际友,刘 球,程 勇,魏志恒,陈明皋,廖德志,黄小飞
(1.中南林业科技大学 林学院,湖南 长沙 410004;2.湖南省林业科学院,湖南 长沙 410004)
干旱胁迫严重影响植物的生长生理进程,成为植物受到的一切非生物损害中,最严重的影响因子之一,对于农林业的生产发展有着直接的影响[1-2]。对于干旱的适应能力,各植物间均存在一定差异,其差异受自身基因调控的影响。植物存在一定的干旱阈值(drought threshold),在其范围内的干旱,会因其抗旱相关基因的表达,产生一系列抗旱性相关的综合性状。作为植物干物质生产的主要途径,光合作用与植物生长密切相关,在干旱胁迫情况下,植物光能容易过剩,而过剩的光能则对植物的器官存在潜在危害。光保护主要是依靠叶黄素循环所产生的热耗散,同时为了阻止光合器官受损,酶促进与非酶促进系统也同时对此进行保护[3-4]。
樟树Cinnamomum camphora为樟科Lauraceae樟属Cinnamomum植物,属于常绿落叶乔木,别名香樟、乌樟、樟木子、芳樟、香蕊[5],喜光喜温,水平分布于10°~30°N 至88°~122°E 之间,通常垂直分布于海拔500 ~600 m 以下的低山平原地区,中国南方的湖南、福建等省份均存在分布[6]。作为我国珍贵的用材树种,樟树有着耐腐朽且防虫害的特点,其纤维长达1 200 ~1 310 μm,密度达0.454 ~0.470 g/cm3,木材呈黄褐色,心材褐黄中带红,樟树同时还是重要的精油原料树种和园林绿化树种,深受我国人民的喜爱[7]。樟树苗期的生长情况会在很大程度上影响樟树后期的生长成材情况。本试验以1年生樟树幼苗为实验材料,通过盆栽控水方法研究不同干旱条件下樟树幼苗的光合特性变化,以便了解樟树幼苗对干旱条件的适应性,为苗期培育过程中的水分管控提供实践与理论指导。
1 材料与方法
1.1 供试材料
选取生长势一致、生长情况良好的1年生樟树实生苗为试验材料,苗木统一使用口径为21 cm的聚乙烯塑料盆栽培,于2017年2月28日上盆栽培。试验场地设于中南林业科技大学生命科学楼B 座顶楼的有机玻璃大棚内,大棚为有机玻璃顶,四面为有机玻璃合围,四面装有窗户且棚内装有通风设施,温湿度与棚外基本一致。
1.2 试验设计
试验采用盆栽控水法人工模拟土壤干旱胁迫,共设置4 个处理梯度,其中正常供水组记为CK,轻度胁迫组记为LS,中度胁迫组记为MS,重度胁迫组记为SS,各组土壤含水量分别为土壤田间最大持水量的90%~85%、75%~70%、55%~50%、35%~30%。每个处理种植3 盆,设置4 个重复,共48 盆。于2017年5月30日开始进行干旱胁迫,连续控水30 d,于每日18:00 对各盆栽进行称质量,补浇当天流失水量,保持土壤湿度在预设值。
1.3 叶绿素含量的测定
叶绿素含量的测定采用张宪政丙酮乙醇混合液浸提法[8],称取0.1 g 叶片均匀剪成1 mm细丝,浸泡于25 mL 丙酮乙醇(V∶V=1 ∶1)混合溶液中遮光浸泡24 h。用分辨率高的分光光度计测定645、652、663 nm 处的光密度值(OD值),计算各叶绿素浓度与比值,每处理重复测定3 次[9]。
1.4 光合参数的测定
胁迫3、6、9、12、30 d 时,利用LI-6400XT便携式光合作用测量系统(LI-COR,USA)测定樟树幼苗各项光合指标。选取叶片位置相同的成熟叶片进行测量,每组测定6 株,每株选取1 片叶片,于8:30 至11:30 测量净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等光合参数,并计算水分利用效率(WUE)[10]。
1.5 数据处理与分析
采用Execl 进行实验数据的整理以及图表绘制,利用SPSS 22.0 软件对数据进行单因素方差分析,并对分析结果进行显著性分析。
2 结果与分析
2.1 干旱胁迫对樟树幼苗叶绿素含量的影响
由图1~4 可知,干旱胁迫下,樟树幼苗叶片组织的叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素含量(Chl)、叶绿素a/b 均存在先上升后下降的趋势,变化的幅度与干旱胁迫的强度有关。各胁迫组叶绿素均于处理6 d 达到最大值,各处理组Chla、Chlb、Chl 与Chla/Chlb 之间均存在显著差异(P<0.05)。此时SS 组樟树幼苗叶片组织的Chla 较之CK 组上升了32.87%,Chlb 较之CK 组上升了14.65%,Chl 较之CK 组上升了30.59%,Chla/Chlb 较之CK 组上升了15.90%。而后,随着胁迫时间的不断增加,樟树幼苗叶片组织叶绿素开始分解,Chla 与Chlb 不断下降,并且下降程度随着干旱胁迫程度的增长而增加,Chla 的积累程度逐渐低于Chlb。处理30 d,各处理组间均存在显著差异(P<0.05),此时LS、MS、SS 组樟树幼苗叶片组织的Chla 较之CK 组下降了31.88%、44.16%、59.70%,Chlb 较之CK 组下降了24.77%、32.88%、44.59%,Chl较之CK 组下降了33.18%、46.36%、59.20%,Chla/Chlb 较 之CK 组 下 降 了9.30%、16.78%、27.14%。
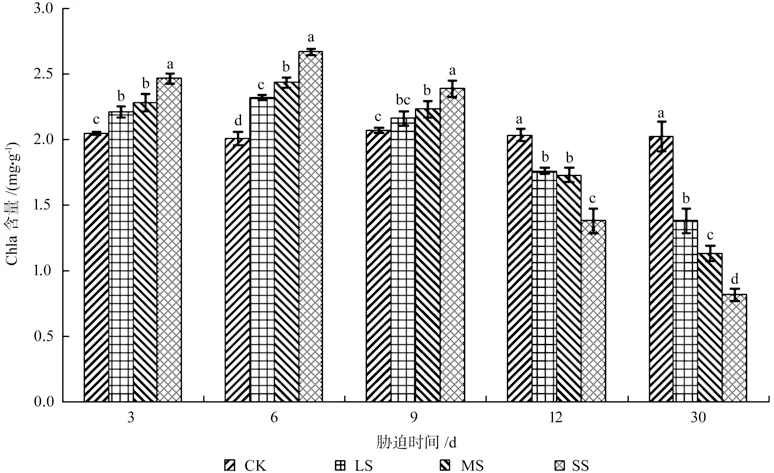
图1 干旱胁迫下樟树幼苗Chla 含量多重比较分析Fig.1 Multiple comparative analysis on Chla content variance of C.camphora seedlings under drought stress

图2 干旱胁迫下樟树幼苗Chlb 含量多重比较分析Fig.2 Multiple comparative analysis on Chlb content variance of C.camphora seedlings under drought stress
2.2 干旱胁迫对樟树幼苗净光合速率的影响
由表1可知,干旱胁迫下樟树幼苗的净光合速率(Pn)随干旱时间的增加呈现下降趋势,且下降的幅度与胁迫强度呈正比,干旱胁迫中胁迫组的净光合速率始终低于对照组。胁迫初期,LS 与MS 组存在一个净光合速率较稳定的时间,LS 组在胁迫9 d 后开始迅速下降,MS 组在胁迫6 d 后迅速下降,此时均与前一时期存在显著差异(P<0.05),说明在轻度胁迫的情况下,樟树幼苗存在自我调控内部生理平衡的能力。处理30 d,各处理组间均存在显著差异(P<0.05),此时LS、MS、SS 组净光合速率较之CK 组分别下降了67.71%、75.95%、90.74%。可见干旱胁迫下樟树幼苗净光合速率受到抑制。
2.3 干旱胁迫对樟树幼苗蒸腾速率的影响
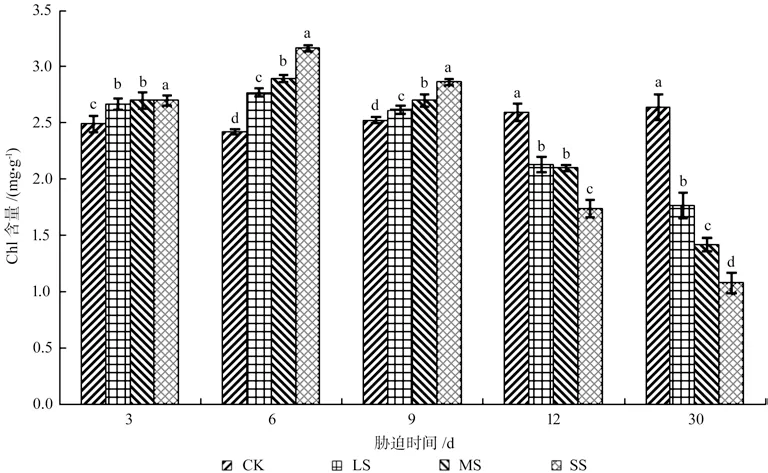
图3 干旱胁迫下樟树幼苗Chl 含量多重比较分析Fig.3 Multiple comparative analysis on Chlb content variance of C.camphora seedlings under drought stress

图4 干旱胁迫下樟树幼苗Chla/Chlb 多重比较分析Fig.4 Multiple comparative analysis on Chla/Chlb variance of C.camphora seedlings under drought stress

表1 干旱胁迫下樟树幼苗Pn多重比较分析†Table1 Multiple comparative analysis on Pn variance of C.camphora seedlings under drought stress
由表2可知,香樟幼苗在干旱胁迫下蒸腾速率(Tr)随干旱时间的延长而降低,且下降的幅度随干旱强度的增加而增加,胁迫期间胁迫组蒸腾速率始终低于对照组。胁迫初期,各胁迫组蒸腾速率均存在下降趋势,处理至6 d,LS 与MS 组间差异不显著(P>0.05),但其他各组间均存在显著差异(P<0.05)。此时LS 与MS 组出现下降的平台期,两组均于处理9 d 急速下降,与处理6 d 时差异显著(P<0.05)。处理9 d,各处理组间均存在极显著差异(P<0.05),直至胁迫结束。处理30 d 时,LS、MS、SS 组较之处理组蒸腾速率分别下降了75.13%、82.72%、90.03%,可见胁迫后期,香樟幼苗的蒸腾速率被大幅度抑制,这是由于土壤干旱导致的叶片气孔关闭的结果,同时也能对植物体的水分流失起到一定程度的控制作用。
2.4 干旱胁迫对樟树幼苗气孔导度的影响
由表3可知,樟树幼苗的气孔导度(Gs)随胁迫时间的延长持续下降,且下降的程度由干旱胁迫强度决定。干旱初期即胁迫3 d,各胁迫组均出现下降的情况,干旱强度越高,下降幅度越大,此时各组间均存在显著差异(P<0.05)。处理9 d,LS 与MS 组的气孔导度均出现迅速下降的情况,均与处理6 d 存在显著差异(P<0.05)。处理30 d 时,各组间均存在显著差异(P<0.05),此时LS、MS、SS 组的气孔导度较之CK 组分别下降了86.46%、92.51%、96.52%,可见干旱胁迫大幅度抑制了樟树幼苗的气孔导度。
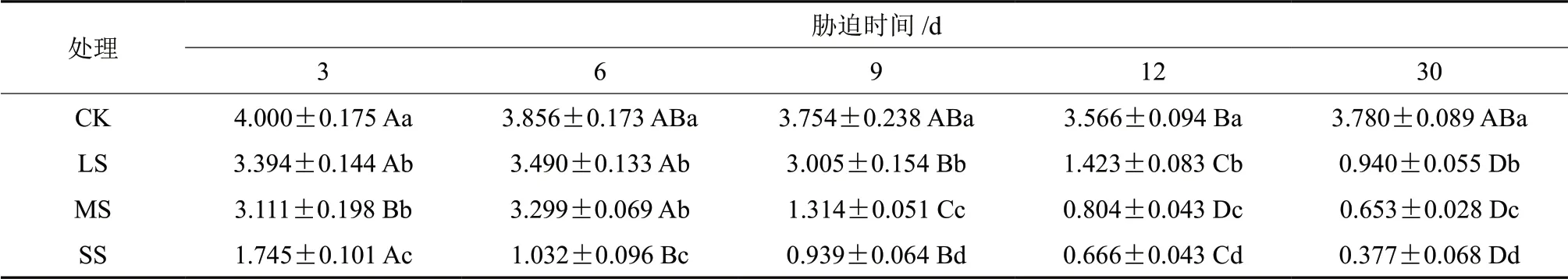
表2 干旱胁迫下樟树幼苗Tr多重比较分析Table2 Multiple comparative analysis of Tr variance of C.camphora seedlings under drought stress
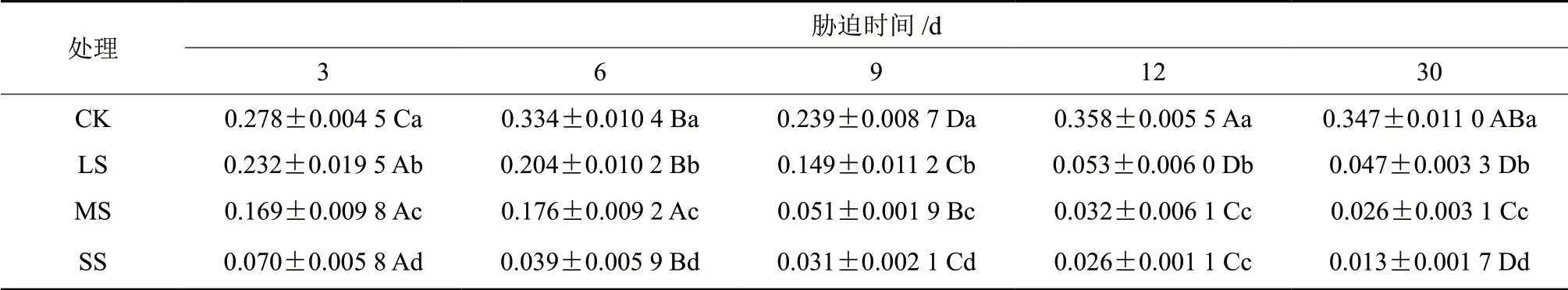
表3 干旱胁迫下樟树幼苗Gs多重比较分析Table3 Multiple comparative analysis of Gs variance of C.camphora seedlings under drought stress
2.5 干旱胁迫对樟树幼苗胞间CO2 浓度的影响
由表4可知,干旱胁迫下樟树幼苗的胞间CO2浓度(Ci)随干旱时间的延长存在下降趋势,且下降的幅度随胁迫程度的加深而加深。轻度干旱与中度干旱组在胁迫初期胞间CO2浓度略微下降后存在平台期,期间下降幅度很小或无下降情况,LS 与MS 组平台期为3 d 到6 d 之间。处理9 d,各处理组间均存在显著差异(P<0.05),此时LS、MS 组平台期结束,胞间CO2浓度开始迅速下降,均与处理6 d 存在显著差异(P<0.05)。处理30 d 时,各组间存在显著差异(P<0.05),此时LS、MS、SS 组的胞间CO2浓度较之CK 组分别下降了49.33%、55.05%、62.27%。
2.6 干旱胁迫对樟树幼苗水分利用效率的影响
由表5可知,随着干旱时间的延长,轻度干旱与中度干旱组水分利用效率(WUE)存在先下降后上升的趋势,而重度干旱组则是先上升后下降,直至低于对照组。处理6 d,LS、MS 组水分利用效率持续下降,与处理 3 d 各组水分利用效率存在显著差异(P<0.05),处理9 d 出现大幅上升,此时MS 组水分利用效率较处理6 d 上升了59.89%并进入稳定,处理12 d 时LS 组水分利用效率较之6 d 时上升了40.20%并进入稳定。处理3 d 时,LS、MS 与CK 组间差异不显著(P>0.05),随着水分利用率的上升,LS、MS与CK 组之间存在显著差异(P<0.05)。SS 组于处理6 d 时达到水分利用效率顶点,为CK 组的60.57%,且与其余各组间存在显著差异(P<0.05),而后持续下降直至处理结束。
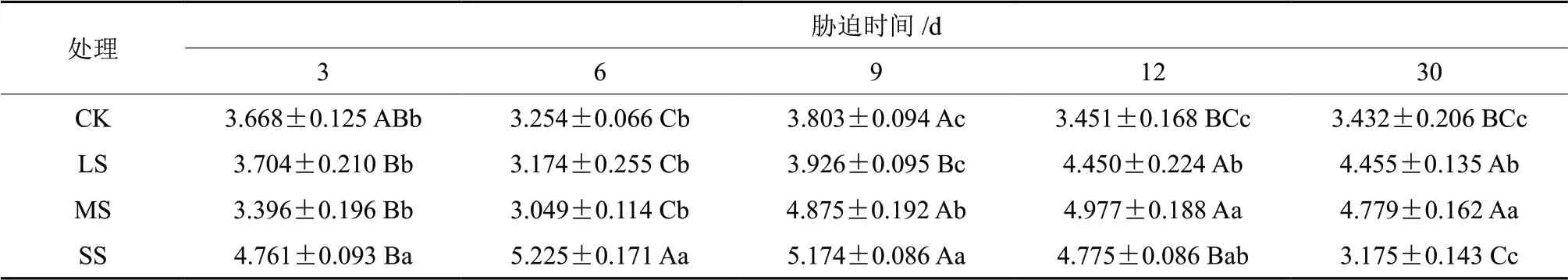
表5 干旱胁迫下樟树幼苗WUE多重比较分析Table5 Multiple comparative analysis of RWUE variance of C.camphora seedlings under drought stress
3 结 论
水分在植物生长过程中扮演着重要地位,是植物健康生长所不可或缺的环境因素,水分的缺乏对于植物的影响是多方面的,且对于光合的抑制作用尤为明显。本试验结果表明,土壤含水量的缺乏严重影响了樟树幼苗的光合生理情况。
干旱胁迫初期,香樟幼苗为了应对干旱条件叶绿素会出现小幅度增加,这是由于植株自身调整导致的应激反应。而随着干旱的时间和程度不断加深,叶绿素的合成受到影响并开始出现分解,使得叶绿素含量随之降低,下降的程度与胁迫的严重程度呈正比。加之香樟幼苗对干旱条件的敏感性下降,Chla/Chlb 的比值也随之降低,说明胁迫中后期香樟幼苗抗旱能力减弱。
在干旱条件下,香樟幼苗植株的光合作用受到抑制,随着干旱时间的延长和干旱程度的加深,净光合速率、蒸腾速率、气孔导度和胞间CO2浓度均存在下降趋势。由于胁迫导致的叶绿素减少,且干旱致使气孔关闭,影响了各项光合生理指标,使得香樟幼苗干物质积累能力受损。而在轻度和中度胁迫的情况下,香樟幼苗的水分利用效率升高,以此抵御缺水影响,而重度胁迫后期水分利用效率下降,说明植株受损严重,自我调控能力下降。
4 讨 论
4.1 干旱胁迫下樟树幼苗叶绿素的变化
干旱胁迫对于樟树幼苗的叶绿素含量、光合情况及水分利用均存在影响,影响的程度与处理有关,各处理间差异显著。叶绿素则是叶片中进行光能吸收、光能传递和光能转换的场所,在植物体内不断进行代谢。植物的光合色素包括叶绿素和胡萝卜素,而其中与植物光合作用关系最密切是叶绿素a,其含量在一定程度上能够体现植物的物质同化能力,进而对植株生长产生影响。植物体内叶绿素在正常状态下能与蛋白质稳定结合,但当植物处于逆境时会导致叶绿素发生不稳定降解。胡义等[11]研究发现,在缺水状态下,由于膜脂过氧化物的作用,植物的叶绿素合成受到抑制,随着干旱程度的加剧,原有叶绿素还会加速分解,导致叶片变得枯黄。研究表明,植物在水分胁迫时,叶绿素会呈先上升后下降的趋势[12-13],这与本试验结果一致。
本试验中,轻度干旱(LS)、中度干旱(MS)、重度干旱(SS)组叶绿素含量均呈下降趋势,且重度干旱(SS)组的下降幅度最大。干旱使得香樟幼苗叶片的相对含水量降低,阻碍了叶绿素的合成同时促进叶绿素分解,使得叶绿素含量随之降低。干旱后期,香樟幼苗对于水分逆境的调节效应减弱,水分利用失衡,导致幼苗对于干旱的抵抗能力逐渐下降。
4.2 干旱胁迫下樟树幼苗光合相关指标的变化
光合作用是植物体进行物质积累的最主要途径,是自然界碳氧循环的主要媒介,植物通过吸收二氧化碳并将碳以生物量的形式固定[14-15]。光保护主要是依靠叶黄素循环所产生的热耗散,同时为了阻止光合器官受损,酶促进与非酶促进系统也同时对此进行保护[16-17]。韩瑞宏等[18]研究表明,紫花苜蓿的叶片在干旱胁迫的条件下,叶片净光合速率、气孔导度、蒸腾速率、叶绿素含量都存在不同程度的下降,叶绿体的超微结构也遭到破坏,随着干旱程度的加深,净光合速率等指标下降相对较慢,叶绿体的外形及其基粒结构也相对变化较小。净光合速率作为在水分胁迫下光合生理中非常重要的一个指标,体现的是单位叶面积的物质生产能力[19],而气孔作为植物与外界水气交流的通道,其密度、数量、大小都会对植物光合作用和蒸腾作用产生重要影响[20]。在干旱胁迫的情况下,植物叶片被迫失水,导致叶片气孔关闭,叶片形态结构受损,对各项光合指标产生直接的影响[21-22]。
实验表明,在水分胁迫的条件下,植物存在适当提高自身水分利用效率的趋势,土壤缺水或根系淹水时,植物的气孔导度和蒸腾速率会随之降低,此时水分利用效率显著提高。但随着胁迫程度的加深,植物叶片生理的动态平衡被打破,净光合速率降低,导致水分利用率也随之降低[23]。本试验中,樟树幼苗在干旱条件下,植株的光合作用受到抑制,随着干旱时间的延长和干旱程度的加深,净光合速率、蒸腾速率、气孔导度和胞间CO2浓度均存在下降趋势。由于胁迫导致的叶绿素减少,且干旱致使气孔关闭,影响了各项光合指标,使得樟树幼苗干物质积累能力受损。而在轻度干旱和中度干旱的情况下,樟树幼苗的水分利用效率升高,以此抵御缺水影响,而重度胁迫后期水分利用效率下降,说明植株受损严重,自我调控能力下降。
本实验仅针对1年生香樟幼苗的干旱胁迫光合生理情况与叶片水分利用情况进行研究讨论,并未结合植物生理生化指标等进行研究分析,研究不够全面。下一步将对香樟幼苗在干旱胁迫下的生理生化指标以及根系指标进行研究,以求建立完善的香樟幼苗抗旱体系,并对香樟的培育与速生提供理论依据。
猜你喜欢
西南农业学报(2021年7期)2021-08-23
作文通讯·高中版(2021年6期)2021-07-14
节水灌溉(2021年5期)2021-05-26
西安交通大学学报(社会科学版)(2021年2期)2021-04-06
农业与技术(2020年12期)2020-07-04
中国农业科技导报(2020年12期)2020-03-15
北方音乐(2019年6期)2019-07-16
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
小小说月刊·下半月(2015年12期)2015-05-14
