
基于COⅠ基因的中国及邻近海域部分笛鲷属鱼类DNA条形码研究
2019-05-10 07:04:40唐楚林王业磷
海洋渔业 2019年2期
唐楚林,肖 林,章 群,周 琪,徐 示,王业磷
(暨南大学生态系,广州 510632)
笛鲷属(Lutjanus)鱼类(下文简称笛鲷)隶属鲈形目 (Perciformes)笛鲷科 (Lutjanidae),据Fishbase(http://www.fishbase.org/)记载该属鱼类全世界约有67种,中国南海有20多种[1]。笛鲷主要分布在世界热带及亚热带海域珊瑚礁或红树林生境,是产地名贵经济鱼类,并具有重要生态服务功能。但近年来,一方面受珊瑚礁和红树林被人为破坏和环境污染影响,其栖息地缩小退化[2];另一方面因其肉质鲜美、经济价值高且具有产卵集群现象而遭受过度捕捞,笛鲷资源现状不容乐观[3]。
准确的物种鉴定是开展海洋生态调查和鱼类资源养护措施制订的前提。由于传统的笛鲷分类主要基于外部形态计测特征和体表斑点条纹特征,不仅高度依赖分类专家个人经验,同时也易受不同发育阶段影响,如形态上笛鲷仔鱼表现出与鮨科(Serranidae)和石鲈科(Haemulidae)等在珊瑚礁生息的其它科鱼类类似的形态特征;不同物种间外部形态特征部分重叠,条纹斑点较为类似且不同发育阶段有变化,采集后会褪色[4],因而需要突破表型分类的局限,建立更有效的笛鲷分类鉴定方法。
DNA条形码[5]通过对一段标准目的基因DNA序列进行PCR扩增和测序分析进行物种鉴定,不受外部形态特征和个体发育阶段的影响。2003年HERBERT等[6]对13 320个物种的 COⅠ基因分析发现,多数物种种内遗传距离小于1%,很少大于2%,遗传距离种间接近或大于种内10倍;2005年 WARD等[7]对澳洲207种鱼类的研究表明,绝大部分鱼类都能被COⅠ基因部分序列有效区分。对鸟类、昆虫等[8-9]越来越多的研究表明,基于COⅠ基因的DNA条形码在动物分类、物种鉴定、隐存种的发掘等方面都具有极其广泛的应用。
目前国内外对于笛鲷的研究主要集中在生理生态[10]和分子系统学研究方面[11-13],DNA条形码仅有VICTOR等[14]鉴定不同发育阶段的巴西笛鲷(L.cyanopterus),王中铎等[1]测序分析国内3省5个地点13种笛鲷13个标本等少数报道。上述研究虽解决了部分笛鲷的分类问题,但仅局限于少数种类与地点。本研究测定了中国沿海19个地点11种笛鲷73个COⅠ基因部分序列,并结合从GenBank下载中国及邻近海域69条同源序列,旨在明确部分种类的分类地位,丰富和补充中国笛鲷属鱼类的COⅠ基因DNA条形码数据库,为种质资源的养护调查提供科学依据。
1 材料与方法
1.1 样品来源及分子实验
本研究测定的勒氏笛鲷(L.russelli)、金焰笛鲷(L.fulviflamma)、奥氏笛鲷(L.ophuysenii)、约氏笛鲷(L.johni)、蓝点笛鲷(L.rivulatus)、五线笛鲷 (L.quinquelineatus)、紫 红 笛 鲷 (L.argentimaculatus)、千年笛鲷(L.sebae)、红鳍笛鲷(L.erythopterus)、马拉巴笛鲷(L.malabaricus)和黄笛鲷(L.lutjanus)等中国11种笛鲷73尾为2004年4月—2014年8月取自中国沿海19个地点,并置于95%乙醇中固定保存,依据Fishbase并参照《中国鱼类系统检索》和《南海鱼类志》进行形态鉴定。样品采集情况和下载的序列信息见表1。
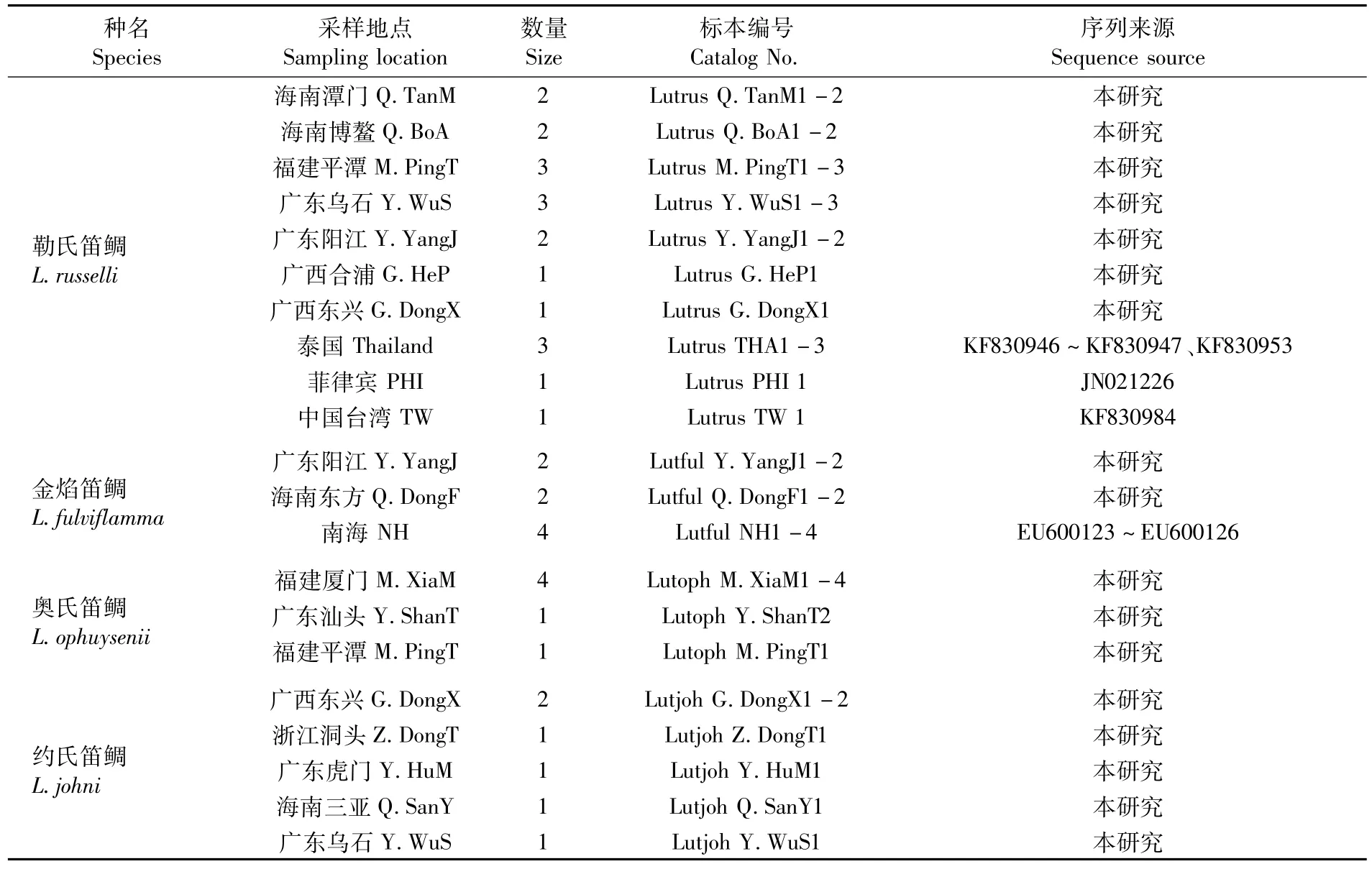
表1 测定或下载的笛鲷属鱼类信息Tab.1 Details of sequenced and downloaded sequences of Lutjanus
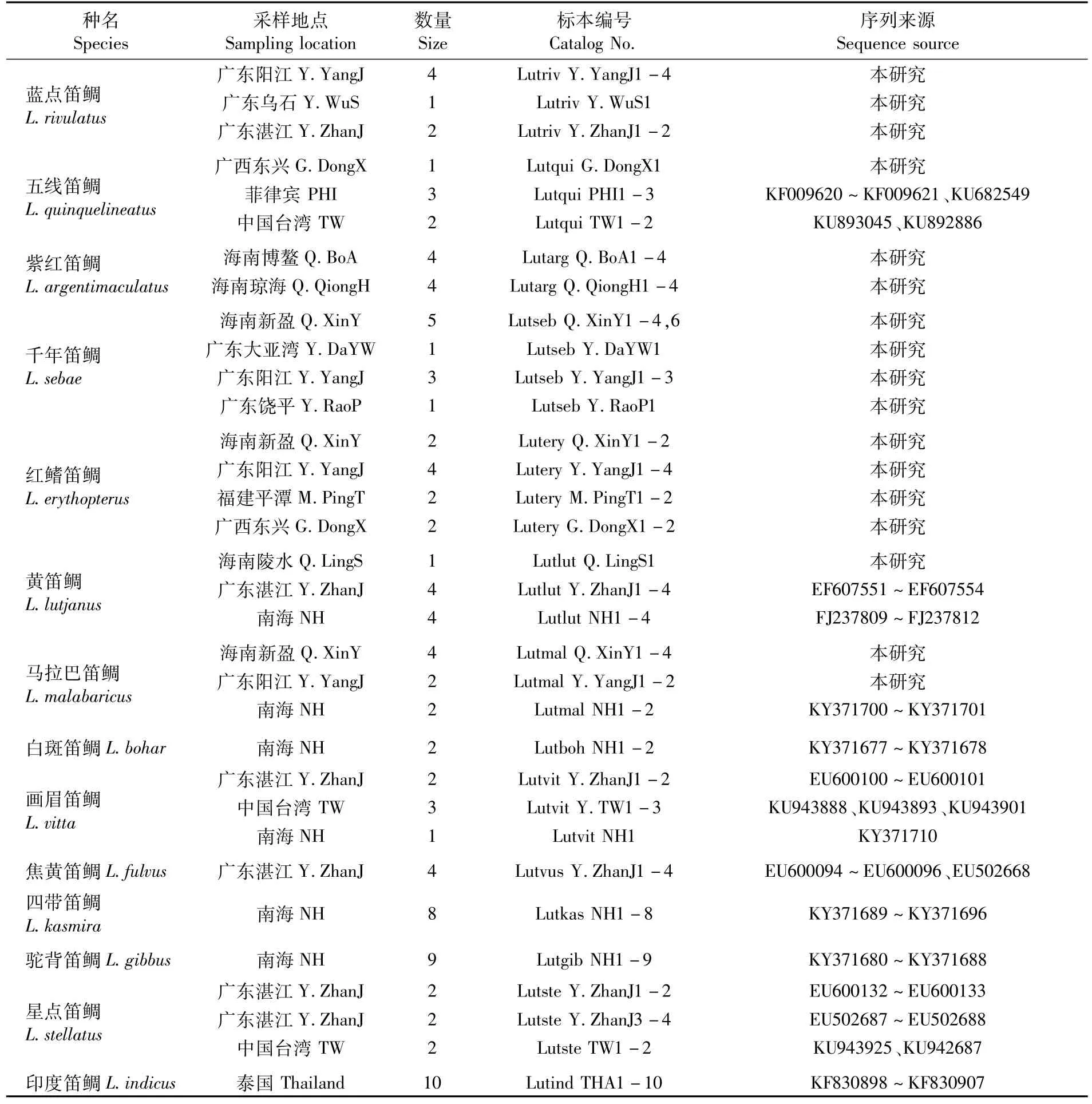
·续表1·
取100 mg鱼背肌肉晾干后,参考乐小亮等[15]改进的酚/氯仿法提取总DNA。使用本实验室自行设计的引物:COⅠF:5′-TGTAAAAC GACGGCCAGTCCTGTGGCAATYACDCGCTGAT,COⅠR:5′-CAGGAAACAGCTATGACNACYTCNG GRTGNCCRAAGAA,采用30μL的 PCR反应体系[16]:上下游引物 (10μmol·L-1)各0.3μL,10×PCR缓冲液 (含1.5 mmol·L-1MgCl2)3μL,BSA和 dNTPs(2 mmol·L-1)各1.2μL,Taq DNA聚合酶(2.5 U·μL-1)0.5μL,DNA模板 1μL,加ddH2O补充至终体积30μL。PCR反应条件为:95℃预变性5 min,95℃变性30 s,55℃退火35 s,72℃延伸45 s,共35个循环,最后72℃延伸10 min。将条带清晰明亮的扩增产物送往北京六合华大基因公司切胶纯化,并于ABI-3730自动测序仪上进行测序。
1.2 数据处理
利用Clustal W软件对测定的序列进行人工校对后,与下载的序列合并,经MEGA7.0软件计算碱基组成、变异位点、简约位点以及转换/颠换的比值,基于Kimura 2-parameter模型(即 K2P)计算种间和种内遗传距离,构建邻接树(neighborjoining tree),并以自举检测(bootstraps)1000次获得支持率[17],同时采用自动条形码间隙检索方法(automatic barcode gap discovery,ABGD)进行假设种分析[18]。
2 结果与分析
截取652 bp的标准条形码序列区[5-7],发现在142条序列中没有碱基插入和缺失;共有223个变异位点,其中210个简约信息位点。碱基T、C、A、G的平均含量分别为 28.1%、28.5%、24.6%和18.8%,A+T含量(52.7%)高于 G+C含量(47.3%),表现出了碱基组成偏倚性,该结果与COⅠ基因碱基组成中普遍存在的AT含量高于GC含量的特征一致[7]。转换与颠换比(R)为3.94,表明碱基突变尚未达到饱和,适合系统发育分析[19]。
基于K2P模型的18种笛鲷的种间和种内遗传距离见表2。结果表明,平均遗传距离种间与种内分别为14.92%和0.55%,种间平均遗传距离是种内的27倍,符合HEBERT[6]提出的10倍种间/种内遗传分歧比的参考标准。种内遗传距离除画眉笛鲷(L.vitta)(2.5%)外,其余均小于2%的种内阈值。种间遗传距离除蓝点笛鲷和星点笛鲷(L.stellatus)间为2.1%、红鳍笛鲷和马拉巴笛鲷间仅有0.3%外,其余物种在4.1% ~21.7%。
在邻接树上,18种笛鲷形成了18个分支(图1),平均遗传距离分支间 14.4% (2.9% ~21.8%)约为分支内 0.17%(0~0.6%)的 85倍(图2),与ABGD分析结果一致(图3):当种内先验遗传距离为0.77%~2.15%时,笛鲷属被分成了18个假设种。18种笛鲷中10种笛鲷独立成支,根据遗传距离种内≤2%,种间≥种内10倍标准[6],支持其物种有效性。另外,勒氏笛鲷进一步分成2个支持率为100%的小支:一支与印度笛鲷(L.indicus)THA1和 THA8混杂在 Clade 1,另一支在Clade 2独立成支;红鳍笛鲷和马拉巴笛鲷聚合在Clade 18上;GenBank下载的画眉笛鲷TW1、星点笛鲷TW1和TW2分别与本实验形态鉴定并测序的奥氏笛鲷和蓝点笛鲷混杂在同一分支上(Clade 6和Clade16)。
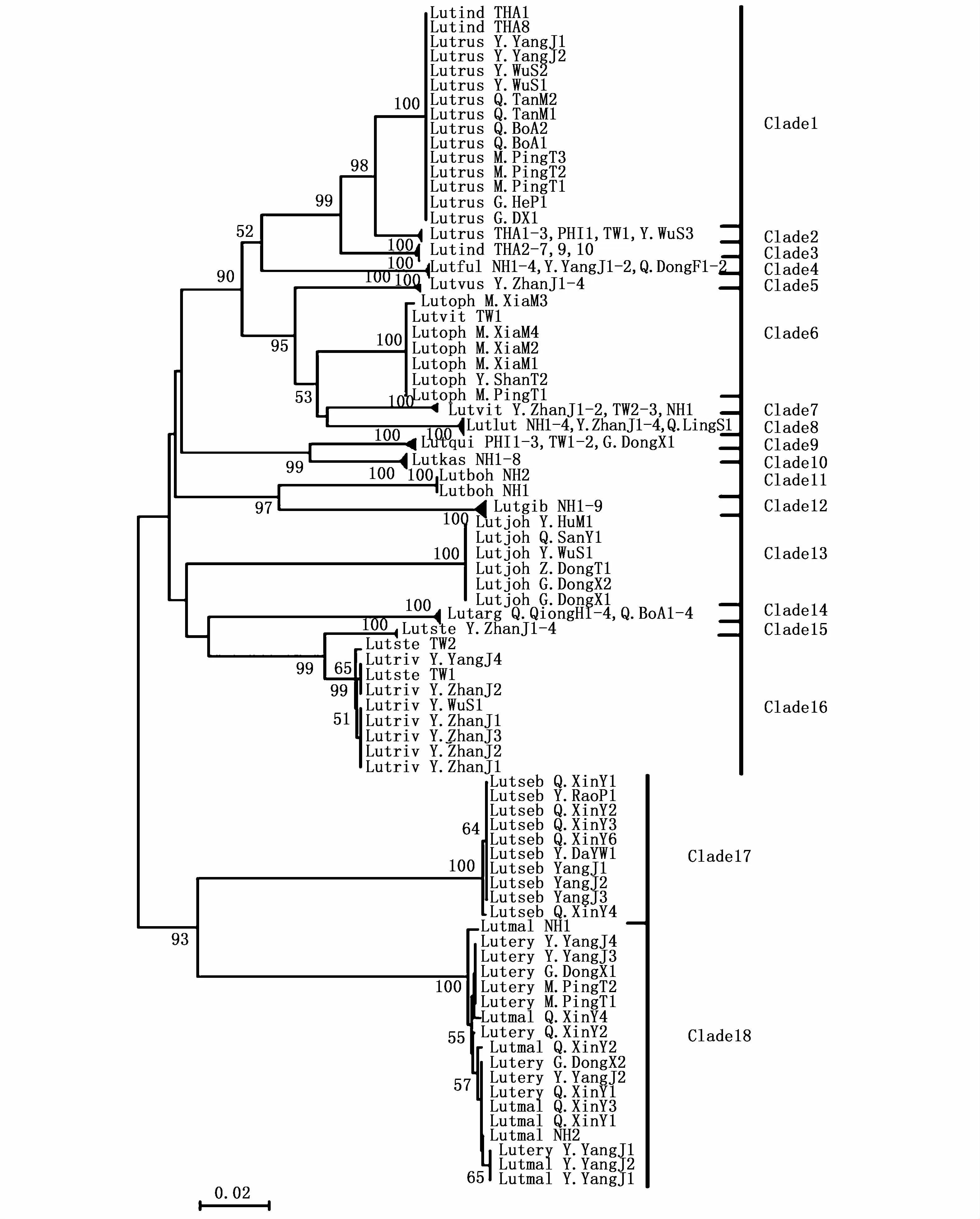
图1 基于线粒体COⅠ基因序列的18种笛鲷邻接树Fig.1 Neighbor-joining tree based on mtDNA COⅠsequences of 18 Lutjanus species
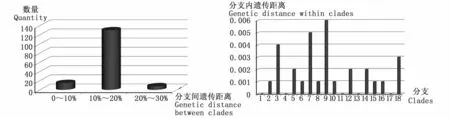
图2 18种笛鲷分支间(左)与分支内(右)遗传距离Fig.2 Histogram of genetic distance between(left)and within(right)clades

图3 笛鲷属鱼类的ABGD分析图Fig.3 Automatic barcode gap discovery analyses of Lutjanus注:横坐标表示先验种内分化值,纵坐标表示划分类群的数量Note:Abscissa means prior intraspecific divergence,ordinate means the number of groups produced
3 讨论
3.1 红鳍笛鲷和马拉巴笛鲷分类地位的探讨
虽然红鳍笛鲷和马拉巴笛鲷体色均为红色,体侧无任何纵带,侧线上方鳞片均斜向后背缘排列,可数性状重叠等形态特征较为相似,但又并不完全相同,如前者侧线下方鳞片均与体轴呈斜行排列,后者侧线下方鳞片则与体轴平行排列,并且鱼尾柄上有黑色鞍状斑且镶以珍珠似的白色边缘[11]。在邻接树上二者聚为一支,分支内遗传距离(0.3%)和种间遗传距离(0.3%)均在一般物种的种内遗传范围(≤2%)内。谭围等[13]也得到类似结果。
这种形态有一定差异但分子上却非常接近的情况,在其它物种中也曾有过报道,如柳淑芬等[20]发现棘头梅童鱼(Collichthys lucidus)和黑鳃梅童鱼(C.niveatus)虽形态有差异,但种间遗传距离仅为0.4%,推断尚未达到物种分化的程度。宫亚运等[21]发现黑斑绯鲤(Upeneus tragula)和马六甲鲱鲤(U.moluccensis)间遗传距离仅有0.3%,但可从体侧纵带和背鳍颜色进行区分,认为可能是近期辐射进化导致的。由此可见,红鳍笛鲷和马拉巴笛鲷可能仍属同一物种,或近期分化的不同物种。另外,由于同属物种种间杂交情况并不鲜见,杂交后代形态上可能倾向于其中一亲本或双亲的中间形态,而线粒体呈母系遗传方式,导致分子与形态鉴定不一致的情况出现,故不能排除种间杂交的可能。
3.2 其它笛鲷分类地位的探讨
1)勒氏笛鲷分为2个100%支持率的小支Clade 1和Clade 2,小支间平均遗传距离(2.9%)是小支内(0.5%)的 5.8倍,种内遗传距离1.3%,推测2个小支可能仍属于同一个物种,但其准确的物种分类地位尚待研究。
2)下载序列星点笛鲷 Lutset TW1和 Lutset TW2与本研究测定序列蓝点笛鲷混杂在Clade 16上,前者种内遗传距离为1.8%,并未达到种的分化水平;且二者的种间遗传距离为2.1%,表明二者可能隶属同一物种,或是没有完成谱系拣选的近期形成的物种,当然也不排除种间杂交或前者错误鉴定的可能。
3)因无法对下载序列进行形态鉴定,对部分类群的分类地位在此仅作如下推测:
①下载的印度笛鲷序列Lutind THA1和Lutind THA8与本实验测序的勒氏笛鲷聚在Clade 1上,分支内遗传距离为0,属种内差异;其余印度笛鲷独立成支。由于印度笛鲷和勒氏笛鲷的外部形态特征较为相似,可能因发育阶段的不同而出现特征重合,推测二者出现混杂可能是鱼类鉴定有误。
②下载的画眉笛鲷序列Lutvit TW1与奥氏笛鲷聚合在Clade 6上,其余的画眉笛鲷占据了Clade 7,分支间遗传距离(6.6%)是分支内遗传距离 0.3%(0.1% ~0.5%)的 22倍,满足HERBERT等[9-10]提出的“10 ×法则”,但画眉笛鲷2个分支间的遗传距离为2.5%,超过种内遗传距离一般小于2%的阈值。从外部形态上看,奥氏笛鲷体侧纵带上有一暗色椭圆大斑,而画眉笛鲷并不具该特征,表明下载的画眉笛鲷序列Lutvit TW1可能隶属奥氏笛鲷。
综上所述,基于COⅠ基因的DNA条形码技术能够对笛鲷属绝大部分鱼类进行有效的区分,说明线粒体COⅠ基因作为笛鲷属DNA分类条形码是可行的,但仍存在一定的局限性,如红鳍笛鲷和马拉巴笛鲷,可能受到错误鉴定、种间杂交或者近期分化等多种因素的影响而出现了聚类在同一分支上的情况,仅进行线粒体COⅠ基因分析不能很好地区分。另外,本研究未能获取中国笛鲷全部报道的物种。因此,在今后的工作中仍需补充其它物种,同时结合形态测量、核基因标记以及生物学研究,以进一步明确中国笛鲷的分类地位。
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
海洋信息技术与应用(2022年1期)2022-06-05 07:38:24
少年文艺·开心阅读作文(2021年8期)2021-09-05 02:57:46
小学科学(学生版)(2019年5期)2019-05-21 01:00:22
少儿美术(快乐历史地理)(2019年11期)2019-04-20 12:33:20
探索科学(2017年5期)2017-06-21 21:16:16
小学生导刊(2017年13期)2017-06-15 20:29:38
金色少年(奇趣科普)(2016年8期)2016-09-21 02:04:58
杂草学报(2015年2期)2016-01-04 14:57:55
