
一株鸽Ⅰ型副粘病毒的分离鉴定及F基因遗传进化分析
2019-04-18 02:32:42杨晓宇恬吕欣孺马沛然杨思远柴洪亮
野生动物学报 2019年2期
李 祥 孙 静 智 敏 杨晓宇 付 恬吕欣孺 马沛然 杨思远 柴洪亮*
(1.东北林业大学野生动物资源学院,哈尔滨,150040;2.黑龙江民族职业学院,哈尔滨,150066)
鸽新城疫(Newcastle disease,ND),又名鸽瘟,是由鸽Ⅰ型副粘病毒(pigeon paramyxovirus serotypeⅠ,PPMV-1)引起的危害禽类的一种急性、高度接触性、烈性传染病。研究认为,PPMV-1是适应宿主鸽子(Columbaspp.)的新城疫病毒(Newcastle disease virus,NDV)变异株[1]。NDV流行范围和宿主范围皆较为广泛,严重危害禽类养殖业,而PPMV-1则是当今严重危害养鸽业的主要病毒性传染病之一。
ND于1923年首先发现于印度尼西亚的巴塔维亚[2],同年在英国新城被发现,由此得名[3]。鸽感染ND最早可追溯到1977年的伊拉克,1981年传播到埃及[4]。随后,1981年意大利的信鸽中检测出了ND[5]。1985年左右,该病迅速扩散,在欧洲、非洲、亚洲和北美洲的多个国家和地区的赛鸽和野生鸽中相继分离出NDV,被认为是ND的第三次全球大流行[6]。
F融合蛋白是促进NDV脂蛋白囊膜与宿主细胞表面包膜相融合的主要因子,F蛋白裂解位点附近碱性氨基酸的数量多少与NDV毒力大小相关,中等毒力和强毒株的裂解位点序列一般为112R/K-R-Q-K/R-R-F117,能够被宿主多种蛋白酶裂解成F1和F2,从而介导病毒粒子与宿主细胞膜融合;而弱毒株只能被胰酶样蛋白酶裂解,一般只引起宿主呼吸道或肠道的局部感染,裂解位点序列为112G/E-K/R-Q-G/E-R-L117。
本研究对东北某动物园中疑似发生Ⅰ型副粘病毒感染的鸽群采集临床样品进行应急检测,从中分离到一株PPMV-1,并对分离株进行F基因序列分析,并通过构建遗传系统发育树确定了其基因型及其与当前鸽源流行株、疫苗株等之间的亲缘关系,为该动物园进行PPMV-1防控提供了有效依据,以期为今后鸽PPMV-1的防控提供理论依据。
1 材料与方法
1.1 实验动物
本试验中9日龄无特定病原体(SPF)鸡胚及SPF鸡红细胞均购自哈尔滨维科生物技术开发公司。
1.2 主要试剂
RNA提取试剂盒购自北京全式金生物技术有限公司;M-MLV反转录酶、RNA酶抑制剂购自Promega公司;PCR相关产品购自北京康润诚业生物科技有限公司;DL 2000 DNA Maker购自大连宝生物工程有限公司。
1.3 引物合成
根据田志革[7]合成NDV鉴定引物,命名为 N1/2,N1:5′-TTGATGGCAGGCCTCTTGC-3′;N2:5′-GGAGGATGTTGGCAGCATT-3′。NDV-F蛋白基因的扩增引物参照熊晓妍等[8]设计,引物序列为 F1:5′-GTCAGTTTACCTGCCTAT-3′;F2:5′-CTCCGACCGTTCTAC-3′。引物由吉林库美生物科技有限公司合成。
1.4 样品采集与病料处理
样品来自于东北某动物园新购入尚在隔离区饲养的2只病死鸽,无菌取鸽的气管、喉头、肺脏、肝脏、脾脏、胰腺、肾脏、直肠,每只鸽的脏器混成2份样品,加入1 mL PBS缓冲液,剪碎研磨,备用。
1.5 病毒分离与鉴定
取0.1—0.2 mL上述研磨上清液由尿囊腔接种9日龄SPF鸡胚,每份样品接种2枚鸡胚,37℃全自动孵化器内孵育72 h,弃去24 h内死胚。无菌环境下收获尿囊液,对收集的尿囊液进行血凝(HA)试验,测定血凝价。
1.6 病毒核酸提取及RT-PCR
按照RNA提取试剂盒说明书要求,对血凝试验结果阳性样品进行总RNA的提取,紧接着用随机引物进行反转录,作为PCR模板。使用NDV通用鉴定引物N1/2进行RT-PCR检测,反应体系如下:模板1.5 μL,双蒸水11.5 μL,2 x PCR Master Mix 10 μL,上游引物和下游引物各1 μL。PCR反应程序:94℃预变性 5 min;94℃变性 30 s,53℃退火 30 s,72℃延伸 30 s,共进行 30 个循环;最后 72℃延伸10 min。使用F1/2引物扩增NDV-F基因,反应体系和程序同上。PCR产物在1%琼脂糖凝胶中进行电泳后,送往吉林库美生物科技有限公司测序。
1.7 序列拼接及遗传进化分析
测序公司返回序列结果用DNAStar软件包中的Seqman程序进行拼接,MegAlign程序进行核苷酸同源性比较。使用Mega 7.0进行序列比对以及氨基酸序列分析,利用IQTREE进行F基因最优进化模型的筛选和最大似然法遗传进化树的构建,并运用Evolview在线工具(http://www.evolgenius.info/evolview)进行进化树的注释和美化。
2 结果与分析
2.1 鸡胚接种及HA试验
以脏器混合研磨上清液接种8枚SPF鸡胚,24 h内死亡2枚,48 h内死亡1枚。对收获尿囊液进行HA试验,所有尿囊液都能使鸡红细胞凝集,效价分别为:24、26、27、27。
2.2 RT-PCR扩增
用NDV通用鉴定引物对分离株核酸进行RT-PCR扩增,反应结果经电泳凝胶检测,结果显示扩增出370 bp左右的特异性条带(图1);选取1株分离株对其命名为pigeon/Northeast China/BS0904/2018(简称BS0904),使用F基因扩增引物扩增,结果显示扩增出1条单一1 800 bp左右特异性条带。条带大小均与预期片段大小相符(图2)。

图1 N1/2引物RT-PCR扩增结果Fig.1 RT-PCR amplification result of N1/2 注:M.DL 2000 DNA Maker;1—4.样品;5.阳性对照;6.阴性对照 Note:M.DL 2000 DNA Maker.1-4.Samples.5.Positive control.6.Negative control

图2 F1/2引物RT-PCR扩增结果Fig.2 RT-PCR amplification result of F1/2 注:M.DL 2000 DNA Maker;1.样品;2.阳性对照;3.阴性对照 Note:M.DL 2000 DNA Maker.1.Sample.2.Positive control.3.Negative
2.3 F基因序列分析
BS0904分离株F基因蛋白质编码区(CDS)长度为1 662 bp,编码553个氨基酸,裂解位点氨基酸序列为112RRQKRF117,具有NDV强毒株的典型分子特征。
同源性分析结果显示(图3),分离株F基因与疫苗株LaSota、B1、Clone 30、VG/GA、V-4、Mukteswar及经典强毒株F48E9的核苷酸同源性为84.2%—85.8%,这说明本分离株与经典的NDV间存在较大差异;而与2011—2013年在中国多个地区鸽中所分离毒株核苷酸同源性则高达98.8%—99.9%,差异较小。
将本研究分离株F基因CDS序列同GenBank中公布的已知基因型的序列构建最大似然树,使用IQTREE中的ModelFinder模块,基于Bayesian Information Criterion(BIC)标准结果筛选出最佳核苷酸替换模型为TIM2+F+I+G4。遗传进化树结果表明分离株与2011—2013年中国多个地区流行的毒株位于同一分支且有很近的遗传进化关系,属于Class Ⅱ,基因型为Ⅵb(图4)。
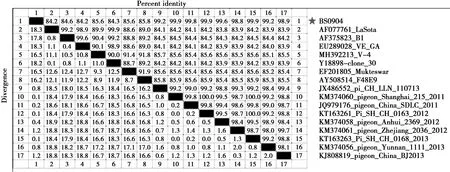
图3 分离株F基因核苷酸序列同源性分析Fig.3 The nucleotide sequence homology analysis of F gene of isolate

图4 基于NDV F基因的遗传进化树及基因型Fig.4 Phylogenetic tree and genotypes of NDVs F gene
3 讨论
鸽新城疫于20世纪70年代末首发于中东地区,1985年在ND第三次全球大流行中传入我国香港,同年8月珠海拱北动物检疫局从引进种鸽中检出该病毒[9],自此以后,该病在我国不断扩散蔓延,目前我国大部分省份都有该病的流行,给养鸽业造成了重大的经济损失。目前我国鸽群中主要流行的毒株为 Class Ⅱ系基因型Ⅵb型鸽新城疫,并且对鸽具有明显的宿主适应性,在系统发育进化树上聚集为一独立分支[10-11]。而基因型Ⅵb型又可分为Ⅵb 1和Ⅵb 2两个分支,其中Ⅵb 1又可分为欧洲早期(EU/re)、欧洲近期(EU/ea)、北美(NA)和伊拉克(IQ)4个亚分支[12]。F基因序列分析结果表明,本研究所分离的BS0904毒株同2011—2013年中国华南和西南多个地区的分离株有着较高的核苷酸同源性(>98.8%),且在遗传进化树上聚集在一起,属于Ⅵb 1-EU/re亚分支,这提示这些毒株具有共同进化起源,这说明此类相关毒株已在中国鸽中有较长时间、较大范围的流行传播。
NDV F蛋白裂解位点氨基酸的序列病毒的毒力有关,F蛋白112—117位氨基酸中,碱性氨基酸越多,致病性往往越强。本研究分离株裂解位点为112RRQKRF117,符合强毒株的典型分子特征。进行疫苗接种是预防鸽群发生ND的有效手段,但免疫失败时有发生,究其原因,除免疫程序不合理、免疫抑制病存在外,疫苗株和流行毒株基因型不一致,抗体不能有效匹配也可能是主要原因之一。BS0904分离株与国内外广泛使用的疫苗株LaSota核苷酸同源性仅为84.2%,与其他疫苗株B1、Clone 30、VG/GA、V-4、Mukteswar同源性为84.2%—85.6%,具有相当大的核苷酸差异。而目前国内鸽群中主要流行的Ⅵb基因型毒株来源并不单一,除了广泛存在的欧洲型外,还有北美型[13]。因此需要针对Ⅵb基因型NDV引发的感染研制新型疫苗。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
科学大观园(2022年2期)2022-01-23 11:05:15
科学(2020年3期)2020-11-26 08:18:22
当代水产(2020年3期)2020-06-15 12:03:02
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
动物医学进展(2015年10期)2015-12-07 05:46:18
医学研究杂志(2015年12期)2015-06-10 06:57:46
特产研究(2014年4期)2014-04-10 12:54:12
