
构树纤维生物酶脱胶过程及影响因素研究
2019-04-10 03:44包振华
生物化工 2019年1期
包振华
(武汉职业技术学院,湖北武汉430074)
构树为桑科构树属落叶乔木,广泛分布于华东、华南、西南,以及河北、山西、陕西、甘肃、湖北、湖南等地[1]。构树皮的韧皮部含有大量的优质纤维,纤维素、半纤维含量高[2]。构树纤维只溶于浓度较高的强酸中,不溶于氢氧化钠、盐酸、二甲苯、二甲基甲酰胺试剂,具有较好的化学稳定性,耐腐蚀性好[3]。
构树纤维具有极高的韧性和极好的吸水吸湿性、透气性,富有弹性,这些性质对其用做纺织、造纸等行业非常重要。
本文从观测研究构树纤维的微观形态及化学组成出发,通过分析构树纤维的排列结构和果胶酶的含量,制备出能对构树纤维有效脱胶的生物酶,研究利用该生物酶对构树纤维进行脱胶及其相应的工艺路线,并分析影响脱胶效果的相关因素。
1 构树纤维的微观形态和化学组成
研究构树纤维微观形态和化学组成的目的主要有两点:一是通过观测构树纤维的长度和其截面形态,计算长细比,看其是否满足作为纺织纤维的要求;二是通过分析构树纤维化学组成中的果胶质含量大小,以此为依据研究制备生物酶脱胶,并探讨相应的技术工艺。
1.1 构树纤维的微观形态
纤维是一种细而长的物质,其直径从几微米到几十微米,长度则从几毫米到几十毫米甚至上千米,长细比很大。一般地,作为纺织纤维,纤维直径≤100μm,长度在几毫米到几十毫米甚至更长,长度/直径>500。
构树纤维的长度一般在4~20 mm,少量纤维的长度大于20 mm或小于4 mm,平均长度约在12 mm。构树纤维平均细度约为0.208 tex,平均强度约为11.515 cN,平均伸长约为6%[3-4]。构树纤维的这种形态结构和相应的物理性能指标适合于作为纺织纤维。
构树纤维作为纺织原料,纤维直径和长细比应能满足要求。在电子显微镜下观察,构树纤维无明显的转曲,表面较为平滑,局部附有少量微细纤维毛以及少量纤维瘤。纤维的胞壁比较薄,厚度比较均匀,内腔空隙较大,两端封闭,两头细,中部粗细较为均匀(图1、2)。构皮中纤维排列紧密,纤维截面呈椭圆形,有压缩感(图3)。

图1 构树纤维纵截面

图2 构树纤维内腔及瘤

图3 构树皮的纤维排列分布
选取10根左右形态较完整的、具有一定代表性的构树纤维,选取20测量点,从构树纤维长度的1/3、1/2、2/3等处测量其直径(图4、5、6)。如表1所示,平均直径约为17.42 μm。
构树纤维直径按20μm计算,纤维长度按10~20 mm计算,则构树纤维长度/直径约在500~1000,符合作为纺织原料的要求。

图4 纤维1/3处
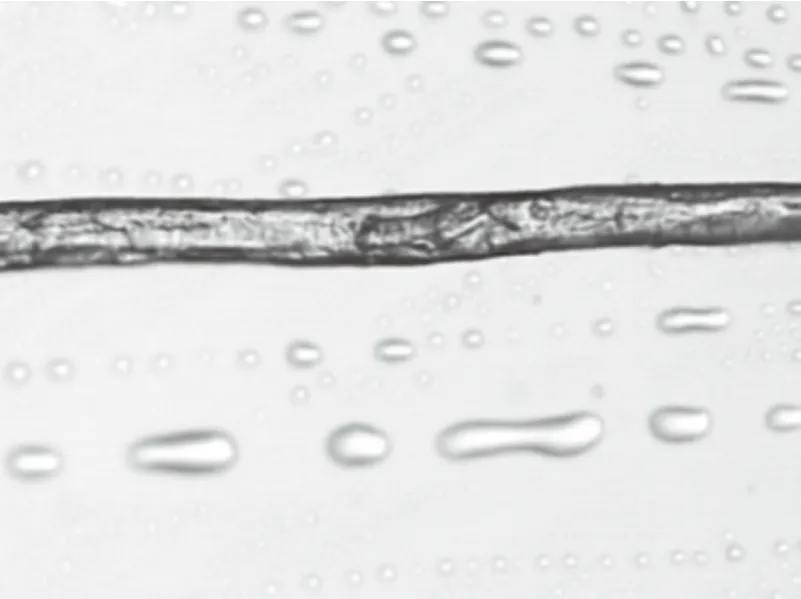
图5 纤维1/2处
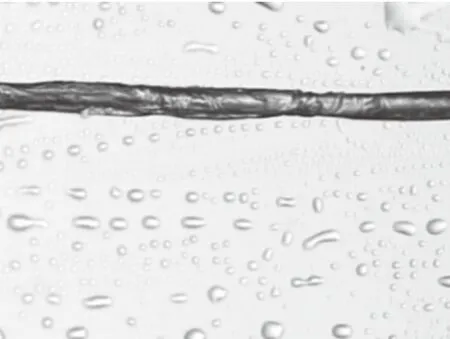
图6 纤维2/3处
1.2 构树纤维的化学组成
研究构树纤维的化学组成,主要是看构树纤维中果胶质含量大小对构树纤维生物酶脱胶的影响。
与其他韧皮纤维一样,构树纤维的主要化学成分为纤维素、半纤维素和木素3种[5]。相比其他韧皮纤维相比,构树纤维的化学比较成分齐全,溶液抽出物的整体比例高,果胶质含量也较高。几种韧皮纤维原料的化学组成比较如表2所示。
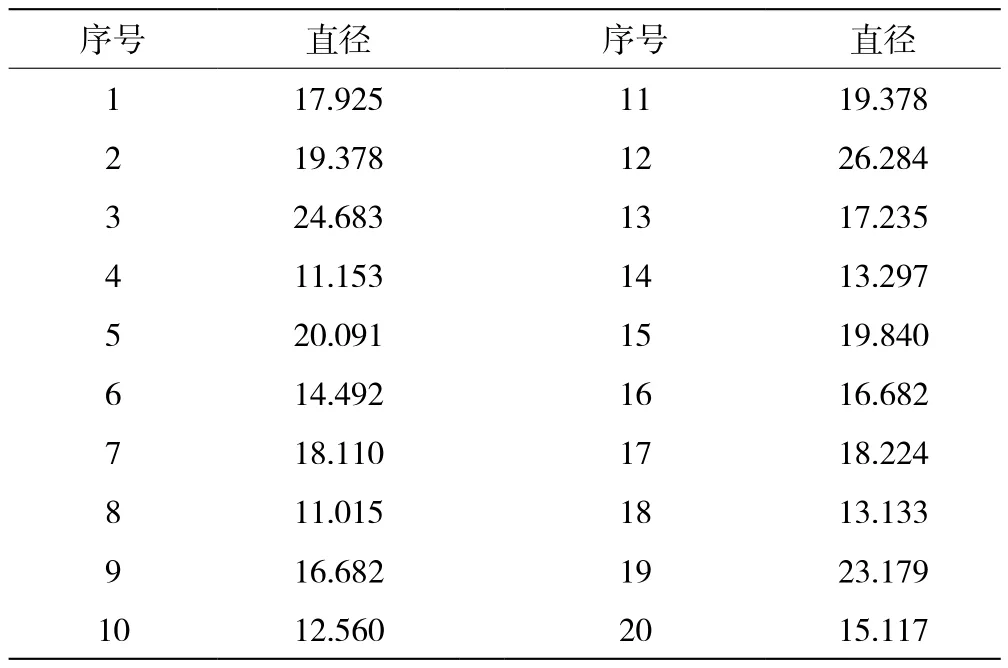
表1 构树纤维的直径测量值(单位:μm)
同是构树纤维,但不同产地的构树皮原料的化学组成略有不同,而且构树全皮与白构皮的化学组成也有一定的差异[6],如表3所示。
从上述两表的构树纤维化学组成中可以看出,构树纤维的果胶含量比其他韧皮纤维高,基本上在10%上下。这一点对研究构树纤维生物酶脱胶非常重要。
2 构树纤维生物酶脱胶技术研究
生物酶是从生物体中产生的,它具有特殊的催化功能。利用生物酶对构树纤维进行脱胶不同于传统的化学方法脱胶,这是由于生物酶具有高效性、专一性、低反应条件以及易变性失活等特点。为使生物酶能对构树纤维进行脱胶,首先是制备能对构树纤维进行脱胶的生物酶。
2.1 生物酶制备
为研究能对构树纤维进行脱胶的生物酶,先查找生物酶生产技术资料,再进行酶菌种筛选。从相关植物中提取酶菌种,用碎玉米粒等做培养基,自然状态下,经过10天左右培育,得到固体颗粒状生物酶初级产品(图7)。再对固体状的初级产品进行细胞壁破碎处理,用盐析法进行简单的分离纯化[7],得到液体生物酶次级产品(图8)。将制得的生物酶溶液在0~4 ℃条件下冷藏备用。
2.2 构树纤维生物酶脱胶基本过程
一般地,构树皮纤维的纤维直径在18~20 μm,以构树白皮厚度1 mm,单纤维直径18 μm来计算,在1 mm厚的构皮中有50~55根单纤维呈平行排列分布,如图3所示。
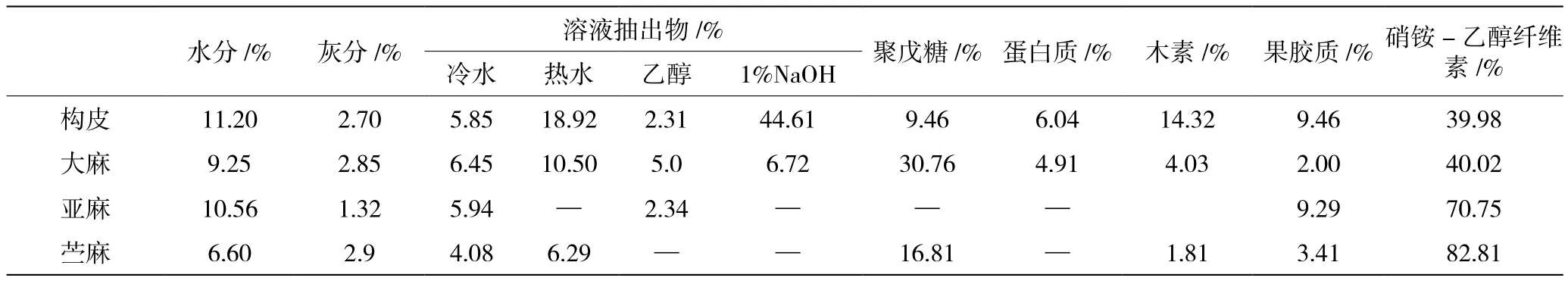
表2 韧皮纤维原料的化学组成
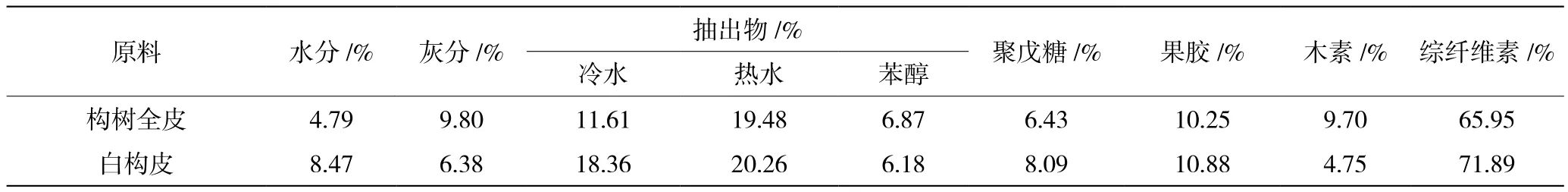
表3 构树皮全皮和白构皮的化学组成

图7 固态生物酶图

图8 液态生物酶图

图9 构树纤维置于生物酶溶液
通过显微观察和分析相关技术参数可以看出,影响构树纤维脱胶质量的主要因素:(1)构树皮中纤维排列过于紧密,脱胶溶液难以快速有效地渗透其中;(2)构树皮中果胶质较高,且成网状分布,将纤维牢固地黏连在一起,不利于纤维脱胶。本文采用自制的生物酶对构树纤维进行脱胶,经过多次反复试验,总体上看脱胶效果良好。
2.2.1 构树皮吸水溶胀
为使生物酶脱胶具有普适性,采用常温条件进行实验。常温下,取刮青后的构树白皮若干,置于容器中,加一定量的自来水,让构树皮完全浸泡其中,放入适量的液体生物酶,静置,观察(图6)。
一般地,经过刮青后的构树皮都会存放一段时间。在含酶溶液中,最初是水从构树皮的表层向里层逐渐溶胀,这就需要的一段时间。水温在26 ℃时,溶胀时间为10~12 h;28℃时,溶胀时间为8~10 h;30 ℃时,溶胀时间在6~8 h;32 ℃时,时间可缩短在6 h左右。
2.2.2 生物酶对构树纤维表层脱胶
构树皮经过一段时间的吸水后逐渐开始溶胀。构树皮在溶胀的同时,生物酶也在不同程度地进行着脱胶作用。由于外层纤维吸水充分,且作用面大,生物酶很容易进行脱胶作用,水溶液中开始出现白色漂浮物,脱胶效果比较明显(图10-1)。这说明生物酶已经开始进行脱胶了。
2.2.3 生物酶对里层纤维进行脱胶
当里层纤维加速溶胀后,生物酶的作用也随之逐渐加快。随着白色漂浮物越来越多,说明脱胶进程越来越快(图10-2)。当构树皮呈完全溶胀状态时,生物酶的脱胶作用越来越明显,纤维间分离速度更快。在自然条件下,水温在26 ℃时,脱胶时间为3~4 d;28 ℃时,脱胶时间2~3 d;30 ℃时,脱胶时间在2~2.5 d;32 ℃时,脱胶时间大约在2 d。
2.3 生物酶脱胶方案
采用生物酶脱胶不如化学脱胶方便快捷。一方面,由于生物酶脱胶作用缓慢,另一方面由于构树皮中纤维排列非常紧密,水溶液不容易渗透其中。为使构树纤维脱胶效果更为理想,可采取两种脱胶方案,即一次脱胶方案和二次脱胶方案。

图10 构树纤维生物酶脱胶过程
2.3.1 一次脱胶
为使脱胶效果良好,在脱胶过程中需要不断翻动构树皮,让其尽可能均匀吸湿膨胀。采取一次脱胶的最大缺点就是脱胶时间较长。
2.3.2 二次脱胶
采用二次脱胶可适当缩短脱胶时间。当纤维溶胀到一定程度、外层脱胶基本完成后,取出构皮,并对构皮进行一定力度的打击,让构皮裂开,再放入容器中进行脱胶。这样既加快了构树皮的溶胀速度,也加快了脱胶速度,脱胶效果更好。
2.4 脱胶后处理
不论是采取一次脱胶还是二次脱胶,构树纤维中都会残留有少量的果胶质。为保证脱胶后纤维干燥时不再粘结在一起,需要将纤维中残留的果胶清除掉。因此,脱胶后还要进行一定的后处理工序。
2.4.1 打纤
当纤维脱胶过程基本完成后,需要对纤维进行适当的捶打,以挤出附着在纤维周围的残留果胶。打纤过程是在构树纤维湿态条件下进行的。利用机械方法对刚脱完胶的构树皮进行反复打击,直到纤维呈单纤维或工艺纤维状。
2.4.2 乳化
经过一定的打击后,极少量的残余果胶还会附着在纤维间,一旦纤维干燥后,这些残余果胶还会将纤维黏连。为此,将经过打击后的离散单纤维放在适量的水中,加入一定比例的乳化液,在100℃条件下煮练15~30 min即可使残留的果胶酶失活,不会再将纤维黏连。
2.4.3 干燥
对构树纤维进行乳化作用后,果胶失活,黏连作用基本消失。再将乳化后的湿态构树纤维进行烘干处理,即可得到离散的构树单纤维或工艺纤维。
3 影响构树纤维生物酶脱胶效果的因素
除了构树纤维的排列结构和果胶质含量外,影响构树纤维生物酶脱胶效果的因素还有纤维原料;生物酶;环境条件以及后处理工艺因素等。这里仅分析几个具有代表性的影响因素。
3.1 构树种类
构树纤维的产地不同、种类不同,构皮纤维中的纤维素及半纤维素含量、果胶质含量等随构树种类的不同而存在一定的差异,这对纤维脱胶有一定影响。
3.2 酶的活性
生物酶的活性与水温呈标准的正态分布。温度过低,生物酶活性会被抑制;随着温度升高,生物酶的活性会逐渐恢复;但温度过高,酶活会被破坏,温度再次降低也不会恢复。
3.3 水溶液的pH值环境
植物纤维中使用的生物酶中大多数最适pH值为6.0~8.0,不同的酶有不同的最适pH值。水溶液最适宜的pH值范围对构树纤维生物脱胶很重要。不同的pH值环境对生物酶活性也有影响。
3.4 生物酶浓度
在一定范围内,生物酶浓度增加,可加快脱胶速度。继续增加生物酶浓度达到一定数值后,脱胶速度将不再增加,而呈现稳定的状态。生物酶浓度不仅影响脱胶效果,也影响脱胶时间和脱胶成本,适当的生物酶浓度既经济又适用。
3.5 打纤质量
韧皮纤维经过生物酶脱胶后,果胶得到不同程度的分解,但由于仍有少量果胶残留在纤维里面,一旦干燥后将重新使纤维发生黏连。为此,需要对纤维进行一定强度的打击。但强度过大会破坏纤维结构,强度过小又起不到打击效果,这就需要适当掌握打击力度。
4 结语
构树纤维具有很多优点,可作为造纸和纺织方面的优质原料。由于构树纤维果胶质含量相对较高,对其有效脱胶尚存在一定的难度。为保护水资源及大气环境,构树纤维脱胶不宜采用化学脱胶或类似的化学脱胶法进行脱胶,利用生物酶技术开发构树纤维非常有必要。
研究生物酶脱胶机理,探索生物酶脱胶技术,实现环保型构树纤维工业化生产,可为造纸和纺织工业提供优质的原料资源。
开发能用于构树纤维脱胶的生物酶,大力开发构树纤维,不仅有助于林业资源的综合开发利用,对生态环境保护和水土保持也具有非常重要的意义。
猜你喜欢
中国麻业科学(2021年5期)2021-12-02
运输经理世界(2021年17期)2021-04-28
粮食与食品工业(2021年1期)2021-02-06
绿色中国(2019年16期)2019-11-26
中国林业产业(2019年3期)2019-03-25
中国扶贫(2017年16期)2017-08-16
西南学林(2016年2期)2016-11-08
中国纤检(2016年8期)2016-10-22
中国麻业科学(2015年5期)2015-12-28
中国造纸(2014年2期)2014-07-24
