
连作对土壤微生物及平邑甜茶幼苗氮吸收、分配和利用的影响
2019-04-09 03:50:30王功帅马子清潘凤兵田长平王金政陈学森尹承苗毛志泉
植物营养与肥料学报 2019年3期
王功帅,马子清,潘凤兵,田长平,陈 杰,王金政,陈学森,尹承苗*,毛志泉*
(1 山东农业大学园艺科学与工程学院/作物生物学国家重点实验室,山东泰安 271018;2 烟台市农业科学研究所,山东烟台 265500;3 临沂市兰山区园林办公室,山东临沂 276000;4 山东省果树研究所,山东泰安 271018)
我国传统优势栽培地区的苹果园主要是上个世纪80年代与90年代初发展建立的,大部分已进入衰老期,其中苹果产区20年龄果园占20%,每年面临2~3.33万 hm2的老果园改造[1],苹果连作障碍是生产中的一大难题,给果农造成巨大的经济损失,严重制约了我国果树产业的可持续发展[2]。土壤pH失衡、自毒类物质、土壤理化性质恶化、微生物因素是引起苹果连作障碍的主要因素[3-4]。苹果连作障碍通常在再植后1~3年内表现,具体表现为地上部发育不良,根系生物量降低,根尖坏死,严重的直接导致植株死亡[5-6]。
连作不仅抑制植物根系的生长,还严重影响对各种元素的吸收和积累。阮维斌等[7]研究表明,连作降低大豆植株对磷、钾等矿质元素的吸收,地上部分的养分比例下降。这可能由于连作提高根系超氧自由基和MDA水平,增加了膜渗透[8-9],过多的自由基使线粒体膜流动性降低[10],膜通透性降低,功能丧失[11]。进而破坏根系皮层和表皮层结构[12],影响根系功能。根系活力和根系质膜ATPase活性是影响根系吸收功能的重要因子。质膜H+-ATPase可利用ATP水解产生的能量将细胞质膜内侧的H+泵到质膜外侧,产生跨膜pH梯度和跨膜电势梯度[13-15]。此电化学梯度为物质跨膜转运提供驱动,带动一系列次级运输体系对各种营养物质、离子的转运[15]。
氮素是果树生长发育过程中所必需的重要营养元素,它影响果树的营养生长和生殖生长,影响果树内各种生理生化过程。氮素的吸收直接关系到器官分化、形成以及树体结构的形成。由于15N标记的特殊性,被广泛应用于植株体内氮素的分配与积累研究上。然而关于苹果连作条件下对氮元素的吸收、利用和分配鲜有报道。本试验以盆栽平邑甜茶为试材,利用15N示踪原子的方法,研究在连作条件下平邑甜茶对15N的吸收、利用和分配,进一步揭示苹果连作障碍的发生机制,为今后克服苹果连作障碍提供理论基础。
1 材料与方法
1.1 试验材料与设计
本试验于2013年1—11月在山东农业大学园艺科学与工程学院国家苹果中心实验站和国家苹果实验室进行。连作土取自山东省泰安市岱岳区满庄镇滩清湾村30年的老苹果园,临近老果园周围的麦田土为正茬。连作土收集距树干1 m、深5—40 cm范围内的土壤,多点取样混匀使用。土壤类型为沙土,有机质含量为5.04 g/kg,土壤硝态氮含量为13.2 mg/kg,铵态氮含量为4.11 mg/kg,速效钾含量为109 mg/kg,速效磷含量为9.28 mg/kg,土壤pH为5.31。正茬土类型为沙土,有机质含量为13.8 mg/kg,土壤硝态氮含量为15.3 mg/kg,铵态氮含量为9.37 mg/kg,速效钾含量为122 mg/kg,速效磷含量为20.4 mg/kg,土壤pH为6.83。
试验材料选用平邑甜茶 (Malus hupehensis Rehd.),2013年1月种子层积,待种子萌动露白后即可在育苗盘中播种育苗。试验采用盆栽的方法,泥盆高26 cm,直径28 cm,装土7 kg,2013年5月选取生长一致的幼苗进行移栽。
本试验设4个处理:T1为连作土溴甲烷灭菌处理;T2为连作土高温灭菌处理;T3为老果园临近麦田土;CK为连作土。每个处理重复20盆共计80盆。待盆栽幼树长势稳定后,于2013年6月3日每盆施用0.5 g15N-尿素 (上海化工院生产,丰度为10.22%),溶于水后均匀浇入盆内。
1.2 测定方法
分别于8月5日和9月5日进行破坏性取样。植株材料用于生物量指标、根系指标和15N的测定;土壤样品用于土壤微生物的测定。每个处理随机选取3个重复。
生物量的测定:平邑甜茶幼苗株高和地径分别用米尺和电子游标卡尺测定,将当天完整取样的幼苗用去离子水冲洗干净,晾干,用天平测定鲜重。将幼苗在105℃下高温杀青30分钟,80℃烘干至恒重为干重。
根形态测定:将不同处理的幼苗根剪下,用清水冲洗干净,铺展在水中,然后用专业版WinRHIZO(2007年版) 根系分析系统对样品图像进行分析处理,记录幼苗根系平均直径、总体积、总表面积和根尖数等根系构型参数。
根系活力的测定参照文献[4]的方法。
根系质膜H+-ATPase的测定参照文献[16]的方法。
土壤微生物的测定:细菌的培养采用牛肉膏蛋白胨培养基,真菌的培养采用马丁氏培养基。
15N的测定:将样品在105℃杀青30 min,80℃烘干,用不锈钢电磨粉碎后过0.25 mm筛,样品装袋暂存待测。15N丰度及全15N含量在中国农业科学院原子能研究所用MAT2251质谱仪测定。
1.3 数据计算与分析
肥料氮 (Ndff%) = (植物样品中15N丰度%-自然丰度%) / (肥料中15N丰度% -自然丰度%) × 100
氮肥利用率 (%) =[Ndff% × 器官全氮量 (g)]/施肥量 (g) × 100
氮肥分配率 (%) = 各器官从氮肥中吸收的氮量(g) / 总吸收氮量 (g) × 100
从氮肥中吸收的氮量 (g) = 器官全氮量 (g) × Ndff%
试验数据采用Microsoft Excel 2003进行计算和作图,通过SPSS 19.0软件进行Duncan显著性检测。
2 结果与分析
2.1 连作对平邑甜茶幼苗生物量的影响
由表1可以看出,连作显著抑制了平邑甜茶幼苗生物量的生长。8月份,连作土溴甲烷熏蒸、高温灭菌与麦田土处理幼苗的生物量没有差异,但显著高于连作土对照处理。连作土对照处理幼苗的鲜重分别为连作土溴甲烷、高温灭菌处理和麦田土处理的56.15%、53.68%、68.22%,干重为溴甲烷、高温灭菌处理和麦田土处理的52.17%、53.33%、66.67%。株高为溴甲烷、高温灭菌处理和麦田土处理的54.77%、59.16%、68.89%。地径为溴甲烷、高温灭菌处理和麦田土处理的80.00%、80.00%、82.35%。9月份,除了地径没有差异外,连作土溴甲烷熏蒸、高温灭菌处理和麦田土处理与连作处理相比幼苗株高、鲜重和干重达到显著性差异。与溴甲烷熏蒸、高温灭菌处理和麦田土处理相比,连作土对照处理的鲜重分别减少了46.77%、46.50%和27.38%;干重分别减少了50.57%、49.04%、39.56%;株高分别减少了41.97%、41.95%和23.51%。
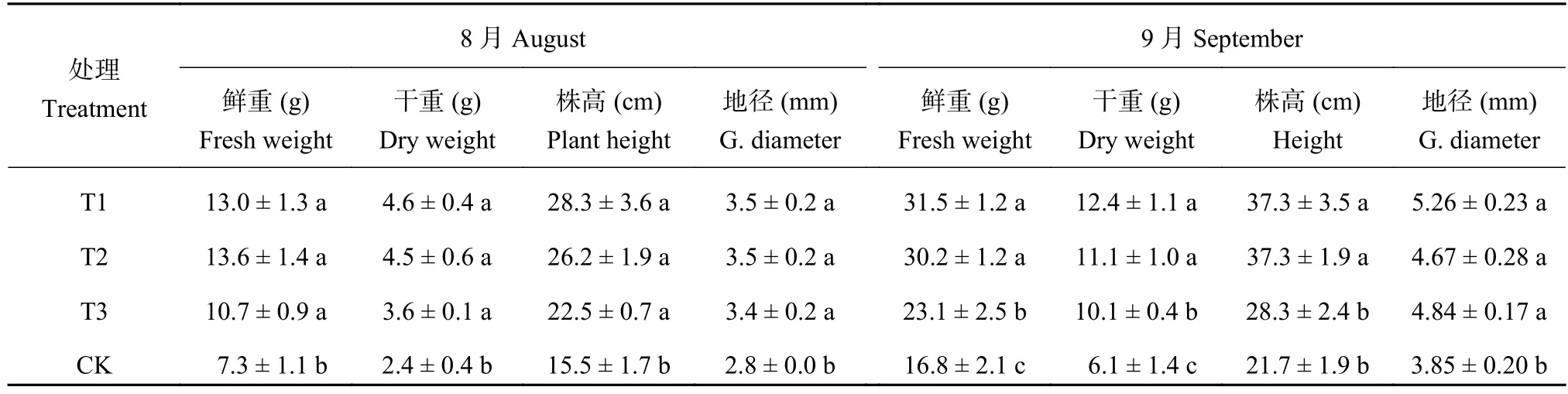
表 1 不同处理平邑甜茶幼苗鲜重、干重及生长状况Table 1 Effect of different treatments on the biomass of Malus hupehensis Rehd. seedlings
2.2 连作对平邑甜茶幼苗根系构型的影响
由表2可知,与连作土处理相比,连作土溴甲烷熏蒸、高温灭菌处理和麦田土处理幼苗根系结构具有明显的差别。连作土对照处理幼苗的根长、根面积、根尖数和分叉数显著低于连作土溴甲烷熏蒸和高温灭菌处理。8月份连作土处理的幼苗根长、根面积、根尖数和分叉数与麦田土处理没有显著性差异。随着植株的生长,9月份测定结果表现为连作土处理的幼苗根长、根面积、根尖数和分叉数显著低于连作土溴甲烷熏蒸、高温灭菌和麦田土处理。与连作土溴甲烷熏蒸、高温灭菌和麦田土处理相比,连作土处理的幼苗根长分别减少了60.10%、59.19%和42.35%;根面积分别减少了56.24%、55.75%和48.07;根尖数分别减少了30.29%、25.78%和25.94%;分叉数分别减少了94.13%、93.87%和93.06%。
2.3 连作对平邑甜茶根系活力的影响
图1显示,连作土处理的幼苗根系活力最低,显著低于连作土溴甲烷熏蒸、高温灭菌和麦田土处理。与连作土溴甲烷熏蒸、高温灭菌和麦田土处理相比,8月连作土处理的幼苗根系活力分别减少了42.65%、43.09%和33.41%。而连作土溴甲烷熏蒸、高温灭菌和麦田土处理无显著差异。9月份麦田土处理幼苗根系活力显著低于连作土溴甲烷熏蒸和高温灭菌处理,高于连作土处理,与连作土溴甲烷熏蒸、高温灭菌和麦田土处理相比,连作土处理的根系活力分别减少了39.71%、40.64%和26.80%。
2.4 连作对平邑甜茶幼苗根系质膜H+-ATP酶的影响
通过测定H+-ATPase,发现连坐土溴甲烷熏蒸和高温灭菌处理幼苗根系质膜 H+-ATP 酶差异不大,但显著高于麦田土和连作土,连作土幼苗根系活性最低,显著低于麦田土。8月和9月趋势基本一致,与溴甲烷熏蒸、高温灭菌和麦田土处理相比,9月份连作土处理幼苗根系的H+-ATPase活性分别减少了41.44%、38.24% 和 25.78%(图2)。
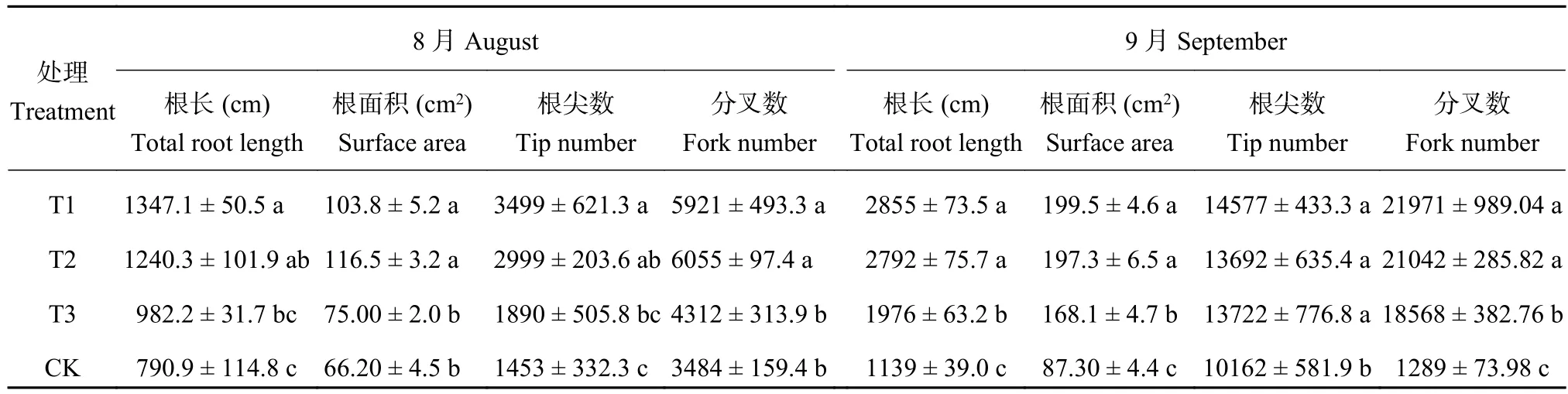
表 2 不同处理对平邑甜茶根系构型的影响Table 2 Effect of different treatments on the root architecture of Malus hupehensis Rehd. seedlings
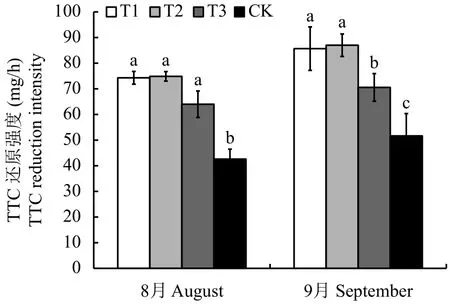
图 1 不同处理对平邑甜茶幼苗根系活力的影响Fig. 1 Effects of different treatments on root activity ofMalus hupehensis Rehd. seedlings
2.5 连作对土壤微生物的影响
由表3可知,不同处理的土壤微生物数量差异很大。8月份,由于连作土溴甲烷熏蒸和高温灭菌,土壤微生物总量显著低于麦田土。连作土中细菌总量最少,显著低于麦田土,真菌总量最高,与麦田土差异不大。随着连作时间的延长,连作土壤微生物发生了显著变化。9月份,连作土细菌总量显著低于溴甲烷熏蒸、高温灭菌和麦田土处理,分别占70.25%、37.13%、78.77%。连作土壤的真菌总量显著高于溴甲烷熏蒸、高温灭菌和麦田土处理,分别是其他3个处理的6.24倍、2.79倍、1.76倍。同时,细菌/真菌比值显著低于溴甲烷熏蒸、高温灭菌和麦田土处理。
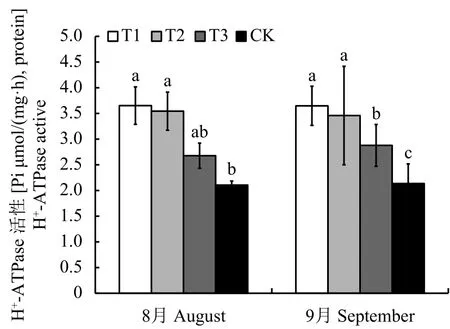
图 2 不同处理对平邑甜茶幼苗根系H+-ATPase的影响Fig. 2 Effect of different treatments on H+-ATPase activeof Malus hupehensis Rehd. seedlings
2.6 连作对15N-尿素的Ndff、分配率和利用率的影响
2.6.1 不同处理对植株各器官Ndff的影响 图3可以看出,连作土处理各器官的Ndff最低,显著低于连作土溴甲烷熏蒸、高温灭菌和麦田土处理。8月份表现为随着植株的生长,各器官的Ndff都增加,但是不同器官的Ndff含量发生了明显变化。连作土溴甲烷熏蒸、高温灭菌和麦田土处理茎叶的Ndff含量高于根系,而连作土处理幼苗根系的Ndff含量高于叶,茎最少。随着15N的不断消耗和植株的生长,在9月份连作土处理根茎叶器官的Ndff与其他3个处理的趋势一致,茎 > 根系 > 叶。连作土处理根系的Ndff含量最低,与连作土溴甲烷熏蒸、高温灭菌和麦田土处理相比,分别减少了18.26%、47.99%和46.78%;茎的Ndff含量分别减少了47.57%、46.75%和45.05%;叶片组织的Ndff含量分别减少了61.34%、58.65%和57.36%。
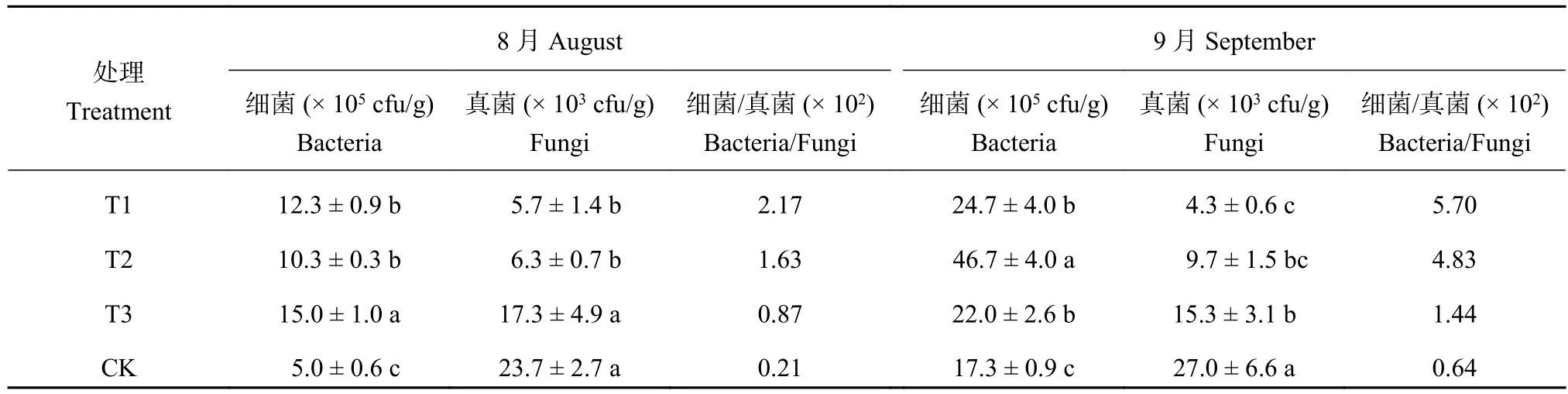
表 3 不同处理对土壤微生物的影响Table 3 Effect of different treatments on soil microbial amount
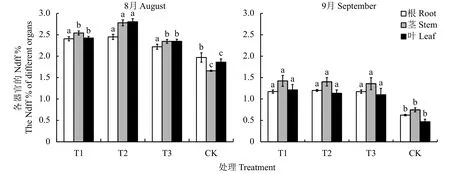
图 3 不同处理对平邑甜茶各器官Ndff的影响Fig. 3 Effect of different treatments on the Ndff in organs of Malus hupehensis Rehd. seedlings
2.6.2 连作对植株各器官15N分配的影响 由图4可知,不同处理平邑甜茶对15N营养元素在不同器官组织的分配有显著的差异。连作土处理的根系和叶组织15N的分配与其他3个处理有显著性差异。连作土幼苗根系15N分配率显著高于连作土溴甲烷熏蒸、高温灭菌和麦田土处理,而叶片15N的分配率显著降低,而茎组织的15N分配基本无差异。8月份连作土处理根系15N的分配率为39.59%,分别是连作土溴甲烷熏蒸、高温灭菌和麦田土的1.16、1.14、1.13倍。随着植株的生长,9月份连作土处理根系15N的分配率有所提高,达到42.11%,是连作土溴甲烷熏蒸、高温灭菌和麦田土处理的1.18倍。叶片组织的15N分配率显著降低,由41.04%减少为29.55%。仅占连作土溴甲烷熏蒸、高温灭菌和麦田土处理的64.78%、73.07%、59.93%。
2.6.3 连作对氮肥利用率的影响 图5可以看出,连作土处理的氮肥利用率最低,显著低于连作土溴甲烷熏蒸、高温灭菌和麦田土处理。8月份连作土处理的氮肥利用率为11.0%,与连作土溴甲烷熏蒸、高温灭菌和麦田土处理相比,分别减少了的61.75%、61.61%和47.68%。9月份连作土的氮肥利用率为13.33%,与其他3个处理相比,连作土处理分别减少了67.19%、67.68%和60.39%。
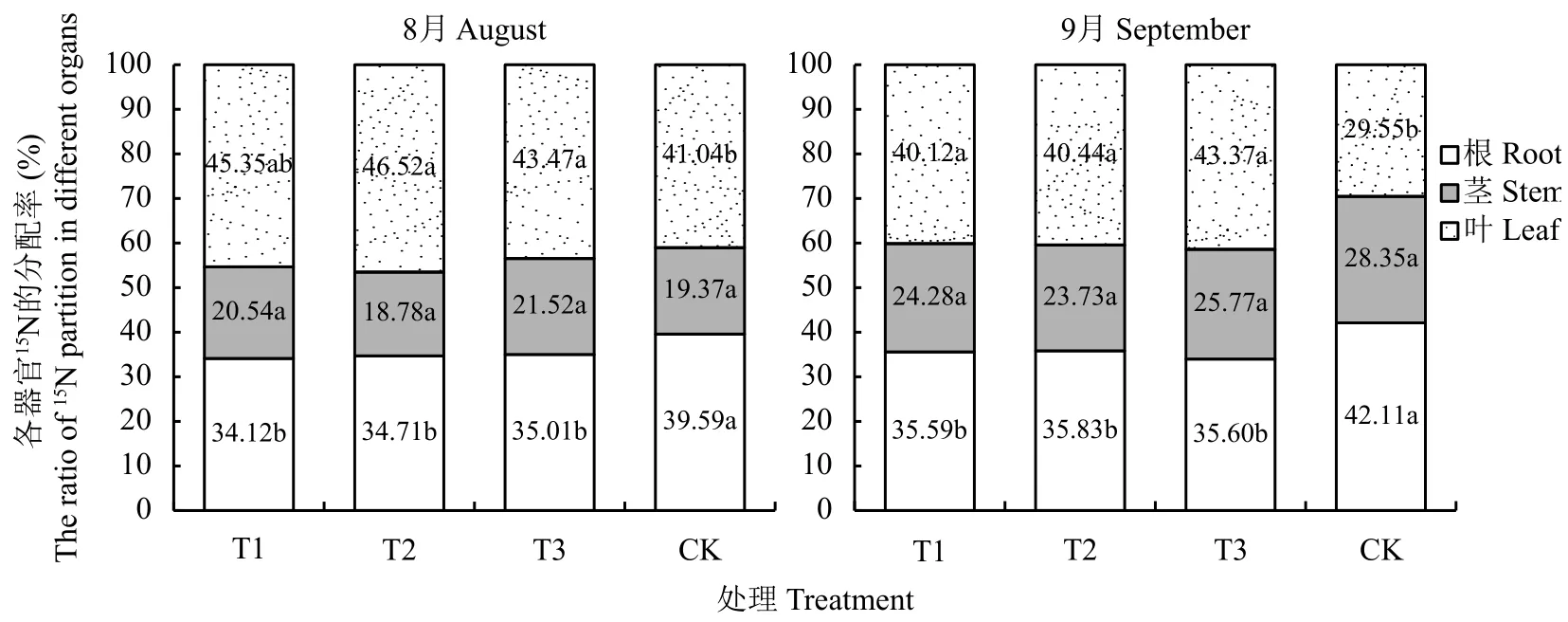
图 4 不同处理对平邑甜茶15N分配的影响Fig. 4 Effect of different treatments on ratio of 15N partition of Malus hupehensis Rehd. seedlings
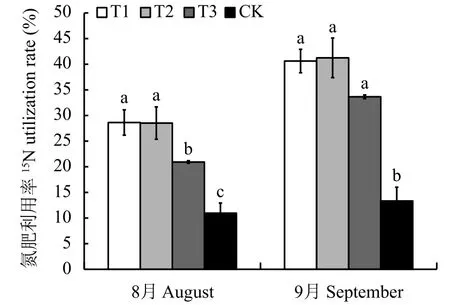
图 5 不同处理对平邑甜茶氮肥利用率的影响Fig. 5 Effect of different treatments on itrogen fertilizer use efficiency of Malus hupehensis Rehd. seedlings
3 讨论
生物因素是引起苹果连作障碍的主要因素[3,17-18]。本试验的结果表明,溴甲烷灭菌和高温灭菌显著提高了连作苹果幼苗的生物量。土壤消毒处理克服了连作障碍的问题,进一步表明有害微生物是引起苹果连作障碍的主要原因,这与Yim等[12]研究结果一致。越来越多的研究表明有害真菌如Phytophthora、Pythium、Cylindrocarpon、Fusarium和Rhizoctonia的增加是引起苹果连作障碍的重要因素[3,18-20]。Franke-Whittle等[21]采用高通量测序方法表明Acremonium、Cylindrocarpon和Fusarium的含量与苹果植株生长呈明显的负相关。病原真菌积累能显著降低农作物产量,增加病虫害[22]。土壤微生物对土壤质量有很大的影响,细菌型土壤是土壤肥力提高的生物学标志之一,真菌型土壤是地力衰退的标志[23]。试验表明,苹果连作显著增加了土壤中真菌的数量,并随着连作时间的延长,土壤真菌数量越多。苹果连作导致土壤由“细菌型”土壤向“真菌型”土壤转变[24]。
根系作为植物吸收水分和养分的主要器官,可以通过其形态和分布来适应外部环境的胁迫[25]。Bauerle等[26]研究表明,根系的快速生长与发达分生能力能够抵御土壤中病原体。本试验结果表明,苹果连作显著抑制了根系的生长,导致根长、根面积、根尖数显著低于正常水平。根系的生长状况是影响果树氮素吸收的关键因素。连作不仅抑制苹果根系生长,还影响根系功能。连作会导致苹果根系自由基大量产生,破坏了抗氧化酶系统和自由基之间的平衡[10,27],过量的ROS会引起质膜氧化损伤,破坏线粒体膜的完整性[27]。连作显著降低了平邑甜茶的根系活力,降低了根系呼吸能力。ATPase是细胞质膜重要的酶,H+-ATPase通过细胞膜运输系统进行跨膜运输增加质子电化学梯度进而提高植物养分的吸收,连作胁迫显著降低了质膜H+-ATPase酶的活性,减少了根系功能所需要的能量,进而影响了膜的功能,降低了根系对营养元素的吸收和运输。
根系吸收的氮素在树体生长过程中起着重要的作用。随着气温的增加和幼苗的快速生长,幼苗对15N的Ndff也不断增加。然而,连作抑制了对15N的征调能力,降低各组织器官的Ndff,影响了不同器官的Ndff的含量。生长初期,连作条件下15N对茎的贡献率最高,叶最低。生长快速期,15N对根系贡献率最高,茎最少。说明更多的氮贡献于根系,减少了植株地上组织生长的供应。这与阮维斌等[7]研究结果一致。同时,连作还影响15N在不同组织器官的分配。试验结果表明根系中的15N分配越来越高,地上部分15N越来越少,并且随时间的延长,根系对15N分配率有增长的趋势。连作引起苹果根系消耗更多的养分,这与Atucha等[28]研究结果一致。苹果对氮肥利用率较低,一般为25%~35%[29]。本试验表明,在8月和9月,连作条件下苹果幼苗的氮肥利用率分别为10.95%和13.33%,显著低于正常水平,连作显著抑制了根系对氮肥利用率。
4 结论
连作抑制了平邑甜茶幼苗根系生长和根系功能,降低了对氮肥的吸收,导致根系过多的消耗营养元素,减少了对地上部分的供应,进而影响植株的生长。因此,在防控苹果连作障碍的措施中,应注重选择抗性和分生能力强的根系,加强根系防护,地上部分增加叶面施肥,缓解连作引起的地上部营养元素供应不足。
猜你喜欢
化学分析计量(2023年7期)2023-08-08 08:59:46
家庭影院技术(2021年7期)2021-08-14 02:58:28
——硫酰氟熏蒸应用技术
粮油食品科技(2021年4期)2021-07-25 03:27:36
生态学报(2019年3期)2019-03-22 01:35:34
中国果业信息(2019年10期)2019-01-05 07:42:30
中成药(2018年9期)2018-10-09 07:19:04
基层中医药(2018年3期)2018-05-31 08:52:10
中国蔬菜(2017年11期)2017-11-13 04:50:28
农产品市场周刊(2016年20期)2016-06-22 16:35:16
现代食品(2016年14期)2016-04-28 08:10:35
