
甜菜碱对低温胁迫下黄芩幼苗生理特性的影响
2019-04-01 01:11李小玲华智锐
贵州农业科学 2019年3期
李小玲, 华智锐
(商洛学院 生物医药与食品工程学院, 陕西 商洛 726000)
黄芩(Scutellariabaicalensis)是我国优质的传统药材,唇形科属,具有安胎止血、清热解毒、泻火燥湿等功效。黄连的抗菌效果在临床应用显著,但黄芩比黄连的药效更好,且不产生抗药性,是一味防治农业病害的理想草药,也是开发潜力较大的药用植物[1]。据史料记载,黄芩始载于《神农本草经》,临床应用已有2000多年的历史,为商洛道地药材,被列入秦岭五大商药之一[2]。近年来,随着中医药现代化的不断发展,全球范围对黄芩的需求量日趋增长,大自然中的黄芩不断锐减,市场对该药材的需求得不到及时补充。因此,人工栽培黄芩成为国内对黄芩药材需求的重要来源。然而当下日渐恶劣的环境严重影响黄芩的产出、产量与质量。商洛地处秦岭山地,昼夜温差较大,春季气温偏低,因此,在黄芩栽培中幼苗常会遭遇冷害,严重影响黄芩的生长[3]。怎样提高植物在低温环境下的抗逆性并获得较高产量是科研工作者长期以来急需解决的问题。近年来,研究发现,植物的生长发育会受到逆境胁迫影响,其中主要制约植物产量和品质的环境因子是低温。植物体内光合作用在低温胁迫条件下受到抑制,损伤细胞膜,细胞老化死亡;细胞内蛋白质降解或变性;基因表达调控改变等[4]。当外界影响引起的损伤超过植物本身的伤害时,植物将表现出冷害症状[5]。因此,探索解决黄芩生产中抗冷性的方法尤为重要。
甜菜碱(Glycine Betain,GB)是一种天然无毒的渗透压调节物质,广泛存在于植物、动物、微生物体内[6]。其作为植物体内一种永久性或半永久性的渗透保护剂,在植物抗逆性中的研究中被广泛关注[7]。在渗透胁迫下,甜菜碱的积累致使许多代谢关键酶保持原来的机能活性,进而保持着细胞膜在低温胁迫下的完整性,保证植株能够正常生长[8]。研究表明,一定浓度下的外源甜菜碱(GB)能提高逆境下植物的保护酶活性,降低丙二醛(MDA)的含量[9-11],提高叶绿素含量[12]。作为药用植物的黄芩,在已有的研究中主要集中在其种质资源、生长发育和育种方法等方面[13]。目前已有外源甜菜碱(GB)对低温胁迫下香蕉幼苗[14]、黄瓜幼苗[15]、结缕草[16]、番茄幼苗[17]等生理特性的影响研究,但有关外源甜菜碱(GB)对黄芩生理特性在低温胁迫下的研究未见报道。且外源甜菜碱在低温胁迫下对植物的研究主要集中在作物、果树、蔬菜等方面,而对药用植物的研究还未见涉及。因此,笔者等以黄芩为试材,研究外源甜菜碱(GB)对黄芩低温胁迫下生理特性的影响,旨在为利用甜菜碱缓解黄芩冷害提供理论依据。
1 材料与方法
1.1 试验材料
商洛黄芩种子:由陕西商洛中药材GAP研究和工程中心提供;外源甜菜碱(GB),购自西安晶博有限公司,含量≥99.5%,分析纯试剂AR级。
1.2 试验方法
1.2.1 材料预培养 选择饱满的黄芩种子于55℃温水中浸泡15 min后捞出,平铺放在培养皿(培养皿上平铺一层滤纸,并湿润)上后,放入恒温箱中催芽。待80%左右的黄芩种子发芽时,播种于50孔穴盘中,放置于人工智能培养箱中育苗。选取长势良好的黄芩幼苗移栽至直径15 cm左右的花盆中进行预培养,共移栽18盆,户外常温,期间定时浇水和松土。预培养30 d后选取生长一致的幼苗用于试验。
1.2.2 试验设计 试验共设2个对照和4个处理。CK1:蒸馏水+常温处理,CK2:蒸馏水+低温处理(4℃)。 根据外源甜菜碱(GB)的浓度设4个处理。T1:0.300 g/L GB,T2:0.600 g/L GB,T3:0.900 g/L GB,T4:1.200 g/L GB。按照以上对照和4个处理,采用不同浓度GB喷施黄芩幼苗叶片,正反面叶片均喷湿。各浓度3次重复,每次重复3株苗,每2 d喷洒1次。处理7 d后,分别选取长势良好的植株放入恒温培养箱中进行4℃低温处理3 d,植株取出后,立即剪下适宜的叶片,洗净,用蒸馏水冲洗,取出主叶脉后,剪碎。用于测定超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、叶绿素含量、过氧化物酶(POD)、可溶性蛋白的含量、丙二醛(MDA)含量和相对电导率等生理指标。
1.2.3 各项生理指标的测定 MDA含量的测定参照文献[18]的硫代巴比妥酸法;SOD活性的测定参照文献[19]氮蓝四唑(NBT)光还原法;POD活性的测定参照文献[20]的愈创木酚显色法;CAT活性测定参照文献[21]紫外光谱法;质膜透性的测定参照文献[21]的相对电导率法;可溶性蛋白含量的测定参照高文献[21]的考马斯亮蓝G-250法;叶绿素含量的测定参照文献[22]丙酮-碳酸钙法。
1.3 数据处理
所有处理每次测定3次重复,数据为3次测定值的平均值,用Excel 2010进行数据统计。统计分析采用SPSS软件,差异显著性用Duncan法检验。
2 结果与分析
2.1 甜菜碱对低温胁迫下黄芩幼苗叶绿素含量的影响
由图1可知,黄芩幼苗叶绿素的含量CK2较CK1低1.698 mg/g,且差异显著,说明低温胁迫降低了黄芩幼苗叶片中叶绿素的含量。当叶面喷施不同浓度GB后,与CK2相比,不同浓度GB处理的叶绿素含量都有不同程度的提高,且随着GB浓度的升高呈先升后降趋势,且T2处理的黄芩幼苗叶绿素含量最高,为3.189 mg/g,较CK2增加66%。由此可知,适宜浓度甜菜碱可提高黄芩在低温胁迫下的叶绿素含量,缓解低温的冷害作用,且喷洒0.6 g/L的甜菜碱效果最佳。
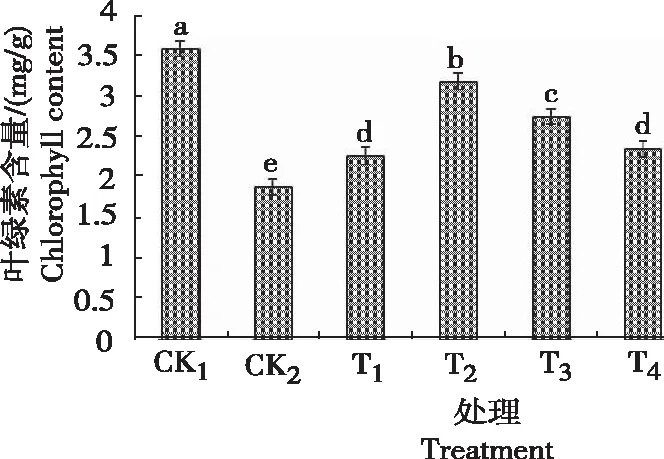
注:图中不同小写字母表示处理间在0.05水平差异显著,下同。Note: Different lowercase letters indicate significance of difference at P<0.05 level. The same below.
2.2 甜菜碱对低温胁迫下黄芩幼苗丙二醛含量的影响
由图2可知,黄芩幼苗叶片丙二醛(MDA)的含量CK2较CK1高9.105 mmol/g,差异显著。且CK2的黄芩幼苗叶片MDA最高,说明低温胁迫加剧了膜脂过氧化作用,引起了大量过氧化产物MDA的积累。当施用不同浓度GB处理后,黄芩幼苗叶片的MDA含量都有不同程度的降低,说明喷施GB能明显抑制MDA生成,且随着GB浓度的升高MDA含量呈先降后升的趋势,T2处理黄芩幼苗叶片的MDA含量最低,与CK2相比降低56%,抑制效果最强。表明,施用甜菜碱可提高黄芩在低温胁迫下的MDA含量,且T2处理(浓度为0.6 g/L的甜菜碱)对黄芩受到低温胁迫影响最小,能有效提高黄芩幼苗的抗逆性。
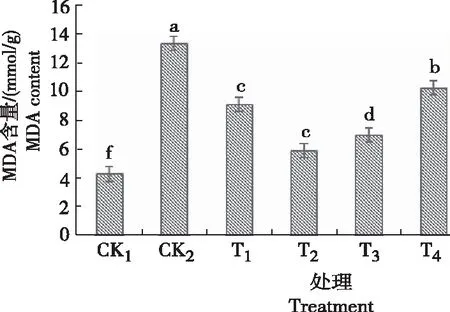
图2 低温胁迫下不同浓度甜菜碱处理黄芩幼苗的MDA含量Fig.2 Leaf MDA content of S. baicalensis seedlings sprayed with different concentration of glycine betaine under low temperature stress
2.3 甜菜碱对低温胁迫下黄芩幼苗SOD、POD和CAT酶活性的影响
由图3可知,CK2与CK1相比,黄芩幼苗叶片中SOD、POD、CAT的活性明显降低,分别降低70.13 U/mg、41.97 U/g·min、6.832 U/g·min,且差异显著,说明低温使植株体内酶的活性大大降低。当施用不同浓度GB处理后,三种酶活力都有不同程度的提升。T1~T4处理后的SOD活力较CK2分别提升了43.23%、50.24%、35.75%和15.38%,且均与CK2差异显著,其中喷施GB为0.6 g/L时效果最佳; T2处理的POD活性最高,比CK2增加26.49 U/(g·min);由图3还可知,低温胁迫下黄芩幼苗叶片中CAT活性先急剧下降,经叶面喷施了不同浓度的外源GB后,CAT活性的变化趋势和SOD、POD趋势基本相同,都有不同程度的提升,总体呈先升后降趋势。T2处理的CAT活性最高,为9.358 U/(g·min),较CK2的3.916 U/(g·min)提高136%。综上所述,喷施GB能够在低温胁迫下提高黄芩幼苗的SOD、POD、CAT酶活力,且效果显著,以T2处理(浓度为0.6 g/L的甜菜碱)效果最佳。
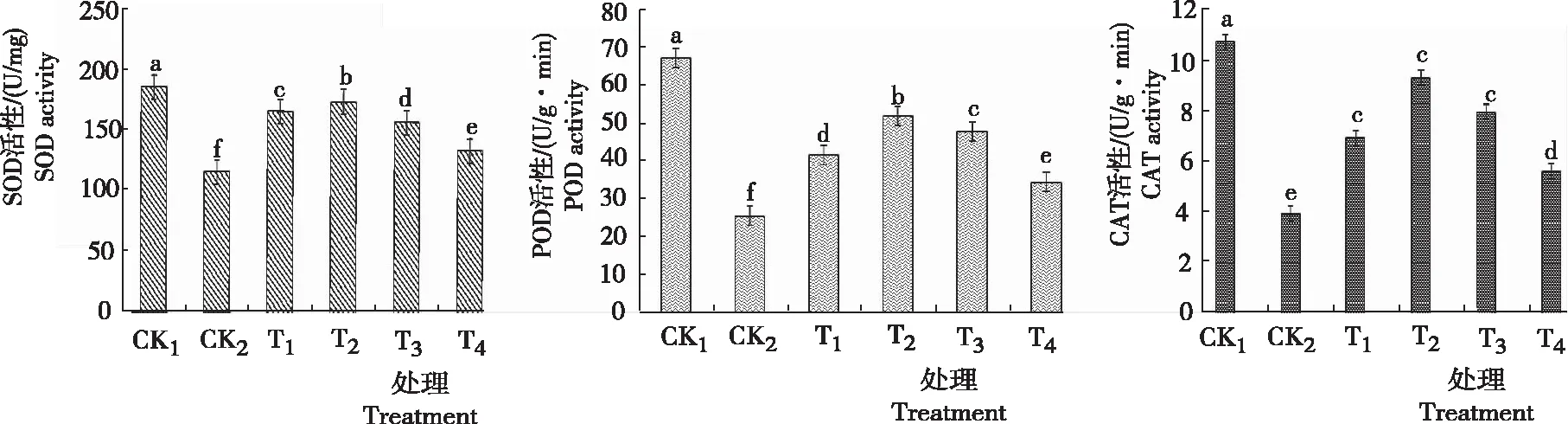
图3 低温胁迫下不同浓度甜菜碱处理黄芩幼苗的SOD、POD和CAT活性Fig.3 Leaf SOD, POD and CAT activity of S. baicalensis seedlings sprayed with different concentration of glycine betaine under low temperature stress
2.4 甜菜碱对低温胁迫下黄芩幼苗可溶性蛋白含量的影响
由图4可知,黄芩幼苗可溶性蛋白含量CK2较CK1降低63.717 mg/g,差异显著,说明低温可使黄芩幼苗的可溶性蛋白含量大大降低。CK2的黄芩幼苗可溶性蛋白含量最低。当施用不同浓度GB处理后黄芩幼苗的可溶性蛋白含量都有不同程度的提升,随着GB浓度的升高,可溶性蛋白含量呈先升后降趋势,且与对照(CK1、CK2)差异明显。T2处理的可溶性蛋白含量最高,为64.146 mg/g,与CK2相比提升178%,说明T2处理(浓度为0.6 g/L的甜菜碱)能显著提升可溶性蛋白的含量。表明,施用甜菜碱可提高可溶性蛋白的含量,能有效提高黄芩幼苗的抗逆性。
2.5 甜菜碱对低温胁迫下黄芩幼苗质膜透性的影响
由图5可知,黄芩幼苗叶片中电导率CK2较CK1显著增加,差异显著,说明低温胁迫导致膜结构破坏,膜透性增大,细胞内容物外渗。CK2与CK1相比,CK2的相对电导率显著增大,比CK1提高154%,说明低温胁迫下质膜透性显著增大。喷施不同浓度GB后,T1~T4处理的相对电导率均有所下降,且随着GB浓度的生高,呈先降后升趋势, T2处理最小,比CK2下降50%左右。表明,喷施GB能明显缓解低温胁迫的迫害。
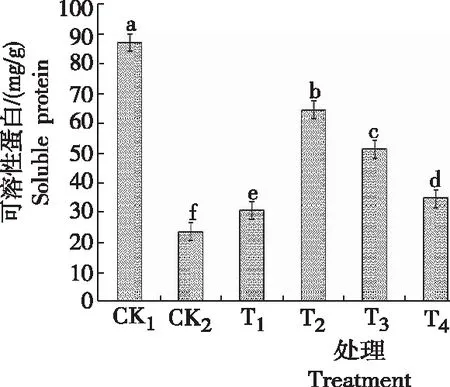
图4低温胁迫下不同浓度甜菜碱处理黄芩幼苗可溶性蛋白的含量
Fig.4 Leaf soluble protein content ofS.baicalensisseedlings sprayed with different concentration of glycine betaine under low temperature stress
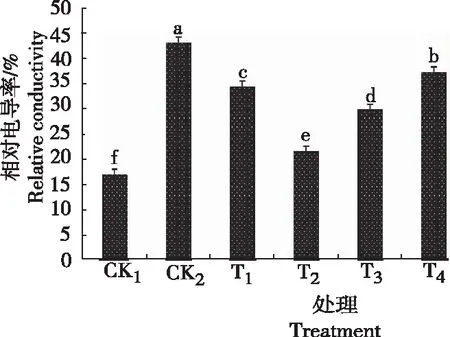
图5 低温胁迫下不同浓度甜菜碱处理黄芩幼苗的相对电导率Fig.5 Leaf relative conductance rate of S. baicalensis seedlings sprayed with different concentration of glycine betaine under low temperature stress
3 结论与讨论
3.1 结论
试验表明,外源甜菜碱可提高黄芩幼苗在低温胁迫下的抗冷能力,适宜浓度的甜菜碱能提高叶绿素含量,促进光合作用;可抑制丙二醛的含量和相对电导率的提高,增加植物细胞内抗氧化系统的活性和抗氧化物质的积累,稳定了细胞膜,继而维持黄芩幼苗的抗冷性。
不同浓度甜菜碱对低温胁迫下黄芩幼苗抗寒能力的影响各不相同,但在浓度为0.6 g/L时,与低温蒸馏水对照相比,黄芩幼苗叶片SOD、POD和CAT活性提高1.502倍、2.031倍和2.362倍;总叶绿素含量提高68%;可溶性蛋白含量高于对照178%;相对电导率和丙二醛含量达到最低,分别下降50%和56%。综合各项指标表明,GB浓度为0.6 g/L时能明显缓解低温胁迫的伤害。
3.2 讨论
大量的研究证明,低温胁迫引起植物的冷害作用是由于植物体内活性氧的积累和酶活性下降所造成的[23],而甜菜碱作为一种天然无毒的渗透压调节物质,除了调节植物细胞的渗透势、维持正常的代谢,还可以提高生物大分子的功能与结构的稳定性,是一类理想的逆境调节物质[24]。适宜浓度的甜菜碱可降低膜脂过氧化产生的丙二醛,提高植物体内的保护酶活性及叶绿素相对含量,有效促进植物光合作用及干物质堆积,从而缓解逆境条件对植物的伤害[25]。
光合作用的强弱与植物本身的叶绿素含量息息相关,植物叶绿素含量下降是植物受到逆境胁迫时的重要特征[26]。卫丹丹[17]认为,低温直接影响叶绿素含量、叶绿体类囊体膜的组分、叶绿素亚显微结构等光合系统的活性和结构。试验中,低温胁迫后黄芩幼苗叶片中叶绿素含量显著下降,说明低温胁迫会对植物光合器官造成伤害。经过甜菜碱的不同浓度处理后,叶绿素含量均有不同程度的提高,且喷施0.6 g/L GB效果最佳,能明显促进叶绿素的合成,进而提高叶片光合活性。
膜脂过氧化作用中,丙二醛作为最重要的产物,通过检测丙二醛的含量,能认识到膜脂过氧化的大小范围,继而间接测定出植物膜系统受伤程度和抗逆性的强弱。所以低温胁迫下MDA含量的高低可以反映黄芩幼苗叶片生物膜受伤害的程度及其抗冷害的强弱。试验表明,低温胁迫下,MDA含量远高于常温正常生长的黄芩幼苗,喷施不同浓度甜菜碱后黄芩幼苗MDA含量均低于蒸馏水+低温(4℃)处理,说明甜菜碱可缓解膜脂过氧化的过程,降低丙二醛的含量,促进植株的正常生长;且喷施0.6 g/L GB能明显缓解低温胁迫的危害。
SOD、POD、CAT是植物体内的保护酶。提高保护酶的活性有利于在低温胁迫下清除大量的活性氧自由基,减少自由基对细胞造成的大分子活性物质的侵害和质膜脂化,从而维持植株各个细胞的正常生命活动[27]。试验表明,在低温胁迫下,施加不同浓度的甜菜碱后,3种保护酶的活性都有不同程度的提高,但都低于常温对照,说明低温影响着保护酶的活性。且以0.6 g/L甜菜碱处理时酶活性提高最大,说明0.6 g/L的甜菜碱能够大幅度提高黄芩幼苗在低温胁迫下的生长指数。
植物抗冷性的增强与蛋白质的合成相关,可溶性蛋白主要起到的作用是维持渗透调节和防止细胞脱水,其含量的增加会提高细胞自身束缚水的含量,从而减少植物脱水的现象的发生[28]。试验表明,在低温胁迫下,黄芩幼苗可溶性蛋白含量大幅度降低,而甜菜碱能够提高黄芩幼苗的可溶性蛋白含量,且在浓度0.6 g/L时,可溶性蛋白含量提高最大,比CK2提高178%。
细胞膜透性大小可通过测量其相对电导率来体现。试验结果表明,在低温胁迫下,黄芩幼苗的相对电导率大幅度增加,经甜菜碱不同浓度处理后,均可显著降低黄芩幼苗的相对电导率;且喷施0.6 g/L GB效果最佳。这可能是由于甜菜碱可以稳定细胞膜上的大分子的结构,保持了酶活性,从而稳定膜结构。
试验结果为黄芩的耐低温性研究提供了参考,也为利用甜菜碱作为化控措施以缓解黄芩冷害提供了理论依据。虽然本研究探讨了甜菜碱在低温胁迫下对黄芩幼苗生理特性的影响,但仅是围绕盆栽试验开展研究,而对于研究黄芩的大田栽培试验中相关生理生化指标还有待继续探讨。
猜你喜欢
现代畜牧科技(2021年8期)2021-10-13
今日农业(2020年16期)2020-12-14
阅读(科学探秘)(2020年8期)2020-11-06
基层中医药(2020年7期)2020-09-11
中国果业信息(2019年1期)2019-01-05
中成药(2017年12期)2018-01-19
生物学教学(2017年9期)2017-08-20
中国洗涤用品工业(2015年8期)2015-02-28
应用化工(2014年9期)2014-08-10
食品工业科技(2014年6期)2014-05-10
