
生物炭-化肥配施对稻田土壤氮磷迁移转化的影响
2019-03-08 02:46王莉霞阎百兴李迎新
农业环境科学学报 2019年2期
崔 虎,王莉霞,欧 洋,阎百兴,韩 露,李迎新,姜 珊
(1.中国科学院湿地生态与环境重点实验室,中国科学院东北地理与农业生态研究所,长春130102;2.中国科学院大学,北京101408;3.吉林大学地球科学学院,长春130061;4.齐齐哈尔大学理学院,黑龙江 齐齐哈尔161006)
生物炭(Biochar)是指生物质在低氧或缺氧的环境条件下,通过控制性高温裂解(400~700 ℃)而产生的一类高度芳香化的难熔性固态高聚产物[1-2]。生物炭具有巨大的比表面积、发达的孔隙结构、较强的吸附能力和丰富的碳含量[3]。同时,也为土壤微生物的栖息和生长提供了良好的环境条件[4]。自然条件下,生物炭呈碱性而被认为是酸性土壤的改良剂[5]。生物炭在高温裂解过程中形成大量羧基官能团,使其表面具有较强的氧化能力和有机质吸附作用,从而使土壤阳离子交换量(CEC)提高53%~538%[6-7]。目前,生物炭在治理农业面源污染、提高耕地质量、应对全球气候变化、维持和稳定农业生态系统功能及保障农业环境安全等方面具有重要意义[8]。
国际生物炭协会(International Biochar Initiative)指出,无机肥减量配施生物炭具有农业经济价值和环境生态效益双重功能[9]。目前,对于生物炭的研究多集中于其制备工艺和理化性质[10]、对污染物环境行为和效应[11]、土壤改良和产量及对某些元素的生物地球化学循环机制的影响[12]等领域。相关研究表明,无机肥减量配施生物炭可降低土壤容重、改善土壤孔隙结构,提高土壤持水能力[13],进而影响土壤氮、磷营养元素的转化过程[14]。有机物包含的氨基酸和氨基糖等含氮物质在高温裂解过程中易凝聚形成杂环氮结构,从而提高土壤有机氮含量,且可通过改变氮素的持留和转化提高土壤氮素有效性[15]。而生物质磷素在热解过程中基本被保留下来,且以可溶态存在,导致土壤有效磷含量显著增加[16]。Brodowski等[17]研究发现,生物炭可富集于土壤微团聚体(<53 μm)中。因而,受益于土壤团聚体的物理保护作用,生物炭中的氮、磷元素有利于长期固持。同时,生物炭因具有较高吸附性能、阳离子交换量和化学反应特性,而常被视为肥料的缓释载体[18],可降低氮、磷养分在土壤中的释放速率,从而降低其淋滤和固定损失量[13]。Ding 等[19]和Laird 等[20]研究发现,生物炭-无机肥配施使土壤-N、-N 和-P 的淋滤量分别降低了15%、11%和69%,从而能够维持农作物生长期土壤的肥力。Mizuta 等[21]和Zeng 等[22]实验证明,无机肥减量配施生物炭(竹木)土壤对上覆水中-N、-N 和-P 的最大吸附量高达17.60、1.25 mg·kg-1和4.96 mg·kg-1,因此,能够降低其随农田退水的流失量。
东北地区作为我国重要的水稻生产基地,对国家粮食安全和保持黑土地生态平衡起着重要作用[23]。目前,对于生物炭配施无机肥在黑土中的应用多集中在不同施用量的对比,而缺乏同等外源添加氮的对比[6,9],因此,较难定量地区分生物炭对于土壤氮磷迁移转化的影响。本研究选取东北黑土区为研究对象,通过田间小区实验,采用生物炭部分替代无机肥的底肥处理方案,在保证外源氮输入量相同的前提下,探讨生物炭-无机肥配施对稻田土壤氮、磷迁移转化的影响,评估生物炭的应用对于土壤和水体氮、磷流失的风险,旨在为生物炭还田管理和农业可持续发展提供科学依据和实践指导。
1 材料与方法
1.1 研究区概况
试验田设置在中国科学院东北地理与农业生态研究所吉林省长春农业综合试验站(125°23′56.30″E,43°59′51.46″N)。该试验站气候类型为北温带大陆性季风气候,土壤类型为黑土,自然条件和供试土壤基本理化性质如表1所示。

表1 试验区自然条件与供试土壤理化性质Table 1 Natural condition of test area and soil physical and chemical properties
1.2 实验设计
以长期耕作的水田为研究对象,在保证各处理外源氮和钾肥输入量相同的前提下,根据生物炭和氮肥不同配施比例,设计4种底肥处理方案(表2),每个处理3 次重复,共计12 个实验小区(4 m×3 m)。各小区均用挡板隔开,以防养分侧渗、窜流。以当地施肥常用类型及用量为依据,底肥选用市售的磷酸氢二铵[(NH4)2HPO4](N 18%;P2O546%;购自吉林福源化肥农药有限公司)和生物炭(原材料:花生壳;pH 8.7;N 0.59%;P2O50.28%;购自河南三利能源有限公司)。水稻种子选用吉粳88,购自长春福田种子有限公司。
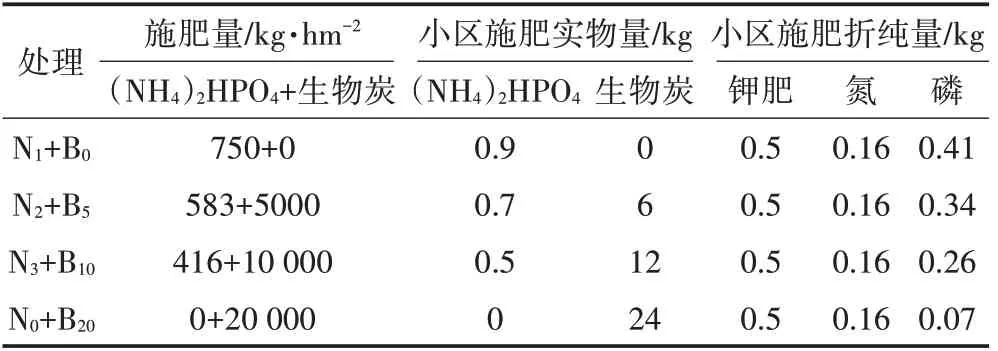
表2 生物炭配施化肥量Table 2 The application amount of fertilizer and biochar
实验起始于2014 年5 月31 日,至2014 年9 月25日结束。稻田各项管理均与本地农户采用的模式相同[24]。具体实施时间如下:5 月31 日,施肥泡田,水位控制在田面以上10 cm;6月5日,初次排水(排水深约8 cm),打浆插秧;6 月15 日,田间除草(丁草胺);7 月20日,追施钾肥(150 kg·hm-2);9月3日,二次排水(排水深约5 cm);9月25日,收割水稻。
1.3 样品采集与分析方法
1.3.1 土样采集
根据水稻不同生长期的养分需求和稻田施肥后土壤氮、磷动态变化特征。于施肥后的第1、10、25、55、85、115 d,采用5 点采样法随机采集稻田表层(0~10 cm)土。自然风干,过100目筛用于测定土样-N、-N、TN 和TP 含量;滴加2 滴甲苯于冷藏(恒温4 ℃)土样中,防止微生物生长,恒温水浴(37 ℃)培养2 h,测定土样脲酶和磷酸酶含量。
1.3.2 水样采集
根据稻田排水时间和施肥后田面水氮、磷动态变化特征,在田面水无扰动的条件下,于施肥后的第2、4、6、10、25、40、55、75、85 d,用医用注射器(100 mL)随机采取田面水(3 次重复),注入聚乙烯塑料瓶,带回实验室。经定量滤纸过滤后,测定水样中TN 和TP含量。
1.3.3 测试方法
1.3.4 数据分析
实验数据采用Excel 2007 进行相关计算;采用SPSS 18.0 进行单指数衰减方程(ExpDec1)拟合和单因素方差分析(One-way NAOVA);采用Origin 9.0 进行绘图。
单指数衰减方程(ExpDec1)是用于模拟指标非稳定变化的数学模型,方程衰减系数(A1)常被视为监测指标浓度衰减速率的表征量。可用于分析稻田土壤中-N、-N、TN 和TP 浓度动态变化趋势,为研究氮、磷素在土壤中的迁移转化规律及土-水界面交换机制提供科学方法。单指数衰减方程:
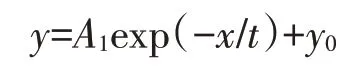
式中:x表示施肥后时间,d;y表示分析指标浓度;A1表示衰减系数(分析指标浓度衰减速率表征量);y0、t 表示单指数衰减方程常数。
2 结果与分析
2.1 生物炭-化肥配施对土壤氮、磷迁移转化的影响
施用不同配比的底肥后,土壤TP 浓度变化差异较为明显(图1d)。N1+B0和N0+B20处理土壤TP 浓度基本呈先略有升高后缓慢下降趋势,分别在0.5~1.5 g·kg-1和0.7~2.2 g·kg-1范围之间变化。N2+B5和N3+B10处理土壤TP 浓度于施肥后第55 d 达到峰值,依次为5.7 g·kg-1和7.1 g·kg-1,之后迅速下降。待土壤TP浓度趋于稳定时,配施生物炭N2+B5、N3+B10和N0+B20处理土壤TP浓度处于同一水平,均为0.7 g·kg-1,且与单施无机肥N1+B0处理无明显差异。综合而言,生物炭、无机肥单施对土壤TP的影响程度较混合配施低。

图1 稻田土壤-N、-N、TN和TP浓度变化Figure 1 Changes in concentrations of -N,-N,TN and TP in paddy soil
2.2 生物炭-化肥配施对稻田土壤和田面水氮磷比的影响
各处理中稻田土壤与田面水氮磷比随时间的变化情况如图2 所示。施肥后第1 d,单施无机肥N1+B0处理和生物炭-无机肥混合配施N2+B5、N3+B10处理土壤氮磷比显著(P<0.05)高于单施生物炭N0+B20处理,分别是N0+B20处理的8.50、7.81 倍和8.19 倍。施肥后第9 d,配施生物炭N2+B5、N3+B10和N0+B20处理土壤氮磷比显著(P<0.05)高于单施无机肥N1+B0处理,分别是N1+B0处理的1.46、1.23 倍和1.19 倍。水稻生长中期和成熟期(25~115 d),4 种处理土壤氮磷比基本呈升高趋势,且配施生物炭N2+B5、N3+B10和N0+B20处理土壤氮磷比均低于单施无机肥N1+B0处理。综合而言,生物炭能够有效调节土壤氮磷比,施肥初期表现为促进作用;而中后期表现为抑制作用。
施肥初期(1~3 d),N2+B5处理田面水氮磷比升高了41.68%。而N3+B10、N0+B20和N1+B0处理田面水氮磷比分别降低了35.41%、32.31%和21.99%。水稻生长旺盛期(6~40 d),4 种处理田面水氮磷比分别在3.20~8.61(N2+B5)、1.42~8.85(N3+B10)、2.35~10.92(N0+B20)和2.14~10.16(N1+B0)范围之间波动。施肥后第55~70 d间(日期:07-25和08-11),N2+B5、N3+B10和N0+B20处理田面水氮磷比显著高于N1+B0处理,分别是N1+B0处理的5.70、3.54、2.73倍和2.37、2.35、8.13倍。水稻成熟期(85 d),4 种处理田面水氮磷比介于8.71~11.10之间。
2.3 生物炭-化肥配施对田面水氮、磷输出负荷的影响
参考三江平原氮、磷输出负荷计算方法[31]。相对单施无机肥N1+B0处理而言,配施生物炭N2+B5、N3+B10和N0+B20处理水田面源污染物-N、-N、TN和TP 的单位面积输出负荷分别降低了42.3%、48.13%、65.24%;55.56%、29.63%、57.41%;39.29%、49.68%、61.69%和38.24%、50.00%、5.88%。表明无机肥减量配施生物炭可有效降低水田面源污染物-N、-N、TN 和TP 的单位面积输出负荷。水田面源污染物-N、-N 和TN 的单位面积输出负荷随生物炭配施量的增加基本呈降低趋势(图3a、图3b、图3c),而TP 的单位面积输出负荷随生物炭配施量的增加呈先下降后上升趋势(图3d)。考虑到肥效和污染物消减总体效果,N3+B10处理是较适合东北黑土区水田的施肥方式。
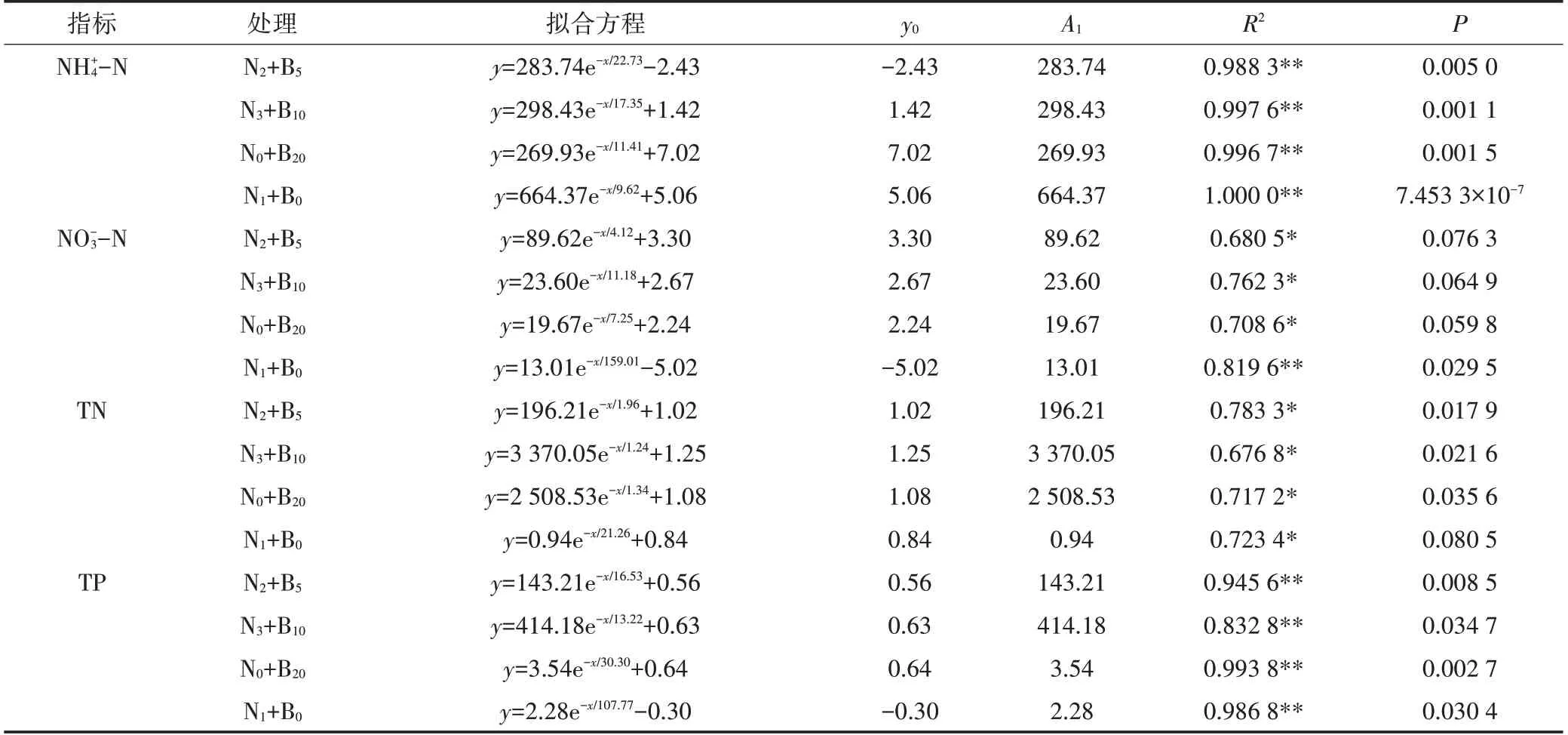
表3 土壤氮、磷浓度单指数衰减方程拟合结果Table 3 The result of N and P concentration change fitted by single exponential decay equation

图2 稻田土壤和田间水氮磷比随施肥时间变化趋势Figure 2 The changes in ratio of TN to TP in paddy soil and field surface water after fertilization

图3 单位面积水田-N、-N、TN和TP输出负荷与生物炭配施量的拟合关系Figure 3 The fitting relationships between -N,-N,TN and TP output load in paddy field and biochar application amount
2.4 生物炭-化肥配施对土壤脲酶、磷酸酶活性的影响
磷酸酶是土壤磷素变化的主导者,可反映其代谢旺盛程度[32]。N0+B20和N3+B10处理土壤磷酸酶活性分别在0.81~0.95 g·kg-1和0.95~1.10 g·kg-1范围之间波动(图4a);N2+B5和N1+B0处理土壤磷酸酶浓度基本呈先下降后上升趋势。水稻生长周期内,配施生物炭N2+B5、N3+B10和N0+B20处理土壤磷酸酶浓度均高于单施无机肥N1+B0处理。表明生物炭施入农田后,可显著提高土壤磷酸酶活性,特别到后期更为明显。
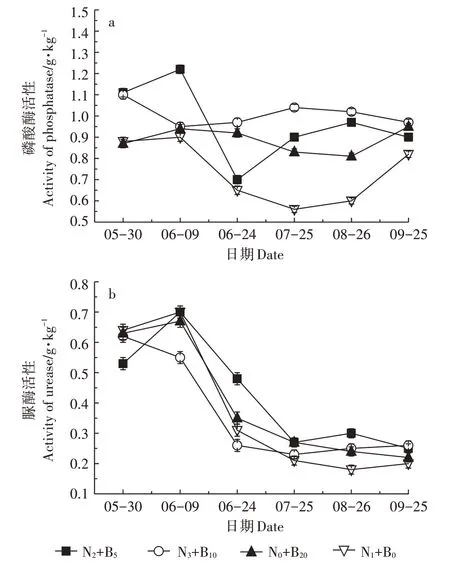
图4 稻田土壤磷酸酶和脲酶活性变化Figure 4 Changes of soil phosphatase and urease activity in paddy soil
脲酶是一种重要的土壤水解酶,其活性与土壤肥力指标显著相关[33]。施肥后25 d 内,N3+B10处理土壤脲酶活性即开始下降,之后基本处于稳定水平(图4b);N1+B0、N2+B5和N0+B20处理土壤脲酶浓度均呈单峰变化趋势,于施肥后第10 d 达到峰值,但浓度差异不明显。水稻成熟期,所有配施生物炭处理土壤脲酶活性均高于单施无机肥处理。因此,生物炭施入农田后可显著提高土壤脲酶活性。
3 讨论
生物炭对土壤中氮、磷元素具有较高的固持容量(表3),从而减弱-N、-N 和-P 在土-水界面的迁移能力,提高肥效的同时,降低其随农田退水流失的风险(图4)。因为生物炭具有发达的孔隙结构[34]、巨大的比表面积[35]和丰富的含氧官能团[36],可通过范德华力与土壤中-N、-N 和-P 发 生等离子交换作用[37],甚至能够通过稳定的化学键对-N、-N 和-P 产 生 不 可 逆 吸 附[35,38]。同时,生物炭小孔隙结构(<0.9 nm)能够降低-N、-N 和-P 的淋溶损失量、延缓水溶性离子的迁移转化时间。Chan等[39]研究发现,无机肥减量配施生物炭使土壤中-N、-N 浓度分别提高了38.0%和4.3%。陈心想等[3]实验证明,生物炭(5~20 t·hm-2)-无机肥配施,土壤TP含量提高了3.8%~38.5%。冯轲等[24]通过田间小区实验,探讨生物炭-无机肥配施对稻田田面水氮、磷流失风险的影响,结果表明,相对单施无机肥而言,配施5000、10 000 kg·hm-2和20 000 kg·hm-2的生物炭田面水-N、-N、TN和TP 的输出负荷分别降低了7.11、8.10、10.98 kg·hm-2;2.70、1.44、2.79 kg·hm-2;10.89、13.77、17.10 kg·hm-2和1.17、1.53、0.18 kg·hm-2。然而,生物炭的施用量对磷的迁移转化能力影响不一,Steiner 等[40]研究发现,高配比(1∶5.52)混施生物炭能够促使土壤中有效性较低的闭蓄态磷向有效磷转化,提高了-P 向田面水迁移的风险。该结论与本研究N0+B20处理结果一致。因此,考虑到肥效和污染物消减的总体效果,N3+B10处理是较适合东北黑土区水田的施肥方式,该施肥方式下水田面源污染物-N、N、TN和TP随农田退水的输出负荷分别降低了29.63%、48.13%、49.68%和50.00%。
生物炭的应用还通过改变土壤酶的活性进而提高土壤肥力和作物产量。土壤酶可作为有机质分解和养分物质循环等生物化学反应的催化剂[41-42],因而被视为土壤生态系统中物质和能量转化联系的纽带。生物炭可通过影响微生物结构变化进而影响土壤各种酶的活性。生物炭丰富的孔隙结构和巨大的比表面积可有效吸附土壤微生物和水溶性有机物,从而为微生物的生长提供了良好的栖息环境[43];生物炭可为微生物的生长提供碳源,且其中的Na、Mo 等微量元素也为微生物的繁殖提供了有利的养分条件[44-45];生物炭可促进土壤团聚体的形成,改善土壤通透性,进而加深土壤颜色,提高土壤温度,增强土壤微生物的新陈代谢能力[46-47]。黄剑[48]研究发现,施用生物炭(4500 kg·hm-2)对土壤酶影响显著(P<0.05)。其中土壤脲酶和碱性磷酸酶活性分别提高了15.6%~248.2%和52.2%~296.0%。周震峰等[49]实验证明,在配施5.0%和1.0%生物炭的条件下,土壤脲酶含量分别是对照的1.10倍和1.52倍。相关研究表明,生物炭可通过改变农田生态系统微生物的丰度和群落结构,进而改善土壤持水性和孔隙度,从而促进植物根系的生长[40]。张伟明等[50]研究发现,生物炭和土壤质量不同配比梯度(生物炭∶土壤=1∶100、1∶150 和1∶200)下,配施生物炭能够增加水稻生育前期根系的主根长、根体积和根鲜重,提高水稻根系总吸收面积和活跃吸收面积,配施生物炭的处理水稻平均增产25.28%。冯轲等[24]通过配施5000 kg·hm-2和10 000 kg·hm-2的生物炭,水稻产量也平均增产了2.06%和1.23%。
土壤氮磷生态化学计量特征是陆地生态系统元素生物地球化学循环偶联的关键指标之一[51]。同时,农田退水营养盐输送也是影响周边水体富营养化的重要原因[52]。因此,研究土壤和田间水的氮磷比可为土壤肥力提供参考[53],也可为防治周边水体污染提供理论指导。水稻生长期内,稻田土壤和田面水氮磷比基本呈先下降后升高趋势。这主要是由于施肥初期生物炭和无机肥中的磷素均以可溶态存在,施入水田后,能够迅速溶解而迁移至田面水。但生物炭中的氮素释放速率相对缓慢,且配施生物炭可提高土壤反硝化细菌的活性,导致土壤-N 在反硝化作用下形成N2或N2O 而挥发散失[54],从而导致土壤氮磷比逐渐降低。生物炭施入稻田后,促使土壤闭蓄态磷向有效态磷转化[40,43]。同时,生物炭能够提高土壤阳离子交换量,增强对Fe3+、Al3+和Ca2+等离子的吸附作用,降低了土壤中磷被固定的风险[55-56]。这可能是导致土壤氮磷比升高的主要原因。Venterink 等[57]研究发现,土壤氮磷比小于14.5 时是氮素受限位点。本研究水稻生长期内,土壤氮磷比介于0.15~1.80 之间,说明氮素是该区域作物生长的主要限制性养分。李如忠等[58]和秦伯强等[59]报道指出,当水体中氮磷比处于10∶1~25∶1 之间时,藻类易呈“爆发性”增长。富氮型(TN/TP>25)和富磷型(TN/TP<10)水体均不利于藻类繁殖。生物炭的添加对于田间水氮磷比的影响不一,水稻生长后期,配施生物炭的处理田间水中氮磷比略有增加,有利于水稻对氮磷的吸收。重点关注的稻田排水期(6 月5 日左右),此时,田面水中氮磷含量较高,而田面水氮磷比介于3.27~10.16 之间(富磷型水体),说明农田退水氮磷比不利于周边水体中藻类繁殖。
4 结论
(2)施肥初期,配施生物炭处理可提高土壤氮磷比,促进水稻生长;水稻成熟期,配施生物炭处理田面水氮磷比显著高于单施无机肥处理,能够持续地给水稻提供营养。
(3)就生态效益而言,N3+B10处理是较适合东北黑土区水田的施肥方式。相对常规施肥而言,该施肥方式下(磷酸氢二铵416 kg·hm-2+生物炭10 000 kg·hm-2),水田面源污染物-N、-N、TN 和TP 的输出负荷分别降低了29.6%、48.1%、49.7%和50.0%。
猜你喜欢
东南大学学报(自然科学版)(2022年4期)2022-08-12
——以长江中游单季稻田为例
湖北农业科学(2022年10期)2022-06-14
山西农业大学学报(自然科学版)(2021年6期)2021-12-28
腐植酸(2021年2期)2021-12-04
腐植酸(2021年2期)2021-12-04
农业与技术(2021年12期)2021-07-02
云南农业(2021年5期)2021-06-19
腐植酸(2020年1期)2020-11-29
农民致富之友(2020年10期)2020-05-11
福建茶叶(2019年12期)2020-01-19
