
连续干旱复水对不同基因型玉米苗期根冠补偿能力的影响
2019-03-08 02:00韩静丽关小康崔静宇丁超明杨明达王怀苹李玉华王同朝
华北农学报 2019年1期
韩静丽,关小康,崔静宇,丁超明,杨明达,刘 影,王怀苹,李玉华,王同朝
(1.河南农业大学 农学院,河南粮食作物协同创新中心,河南 郑州 450002;2.浚县丰黎种业有限公司,河南 浚县 456250)
玉米是我国乃至世界重要的粮食、饲料和经济作物。在黄淮海夏玉米区,玉米生育期内经常因为降雨不足、不均或需水期与降雨期错位而面临土壤干湿交替的多变水分环境条件。在水分匮乏的环境里,玉米常通过减少叶面积、降低光合合成能力抑制代谢作用等维持生长,此类机制均以牺牲干物质积累量或产量为代价,但干旱胁迫对作物的影响并不总是有害的,在不超过其生理响应阈值的范围内进行适度干旱胁迫,然后恢复水分供应,可在生理和产量上形成 “补偿效应”,在节约用水的同时,可获得较高产量并提高水分利用效率[1]。早在20世纪90年代中后期,苏珮等[2]在盆栽玉米、梁宗锁等[3]对分根交替胁迫下的玉米以及Norwood[4]在碗柱草属植物中均观察到干旱后复水条件下植物的生长发育、籽粒产量和生理功能等方面存在补偿效应,即植物经历干湿交替的变水条件后能促进其生长和代谢功能产生补偿效应。
补偿或超补偿其实是植物在漫长的进化过程中形成的一种对水分亏缺环境的适应性,其根本原因在于植物在器官、细胞中的酶组分甚至基因结构方面均存在大量冗余,冗余的存在有助于作物适应环境因子的波动性,但在适宜生长条件下作物的生长冗余一方面消耗物质、能量、时间和空间资源,降低经济产量;另一方面已产生的冗余还会激化植株个体或不同器官间的竞争,产生新的冗余,使得有效资源被浪费,生长冗余就成了负担[5]。适度减小冗余可使植物对获取的有限物质和能量进行最合理、最优化的分配和利用,产生最大超补偿[6],这就是补偿的生态学机制。因此,只要合理设置干旱复水的程度、时期和频次,就可以利用植株在干旱胁迫下的自我保护机制比如老叶程序性死亡、着力新叶生长以及复水后的物质重新分配等生理过程减少生长冗余,实现地上部的超补偿生长,在提高水分利用效率的同时,保持或提高产量[7]。
作物产量的形成是根系与地上部在群体条件下共同作用的结果,而根系早已被人们所重视,其中在植物根源信号是重要的研究热点,早在1985年Blackman等[8]就提出了根冠通讯学说。但之后对根冠通讯的研究大多集中于根冠通讯的机制及信号转导上,忽略了其对根冠间资源分配的影响[9-10]。因此,拟利用玉米苗期盆栽试验,研究连续干旱复水下根冠通讯对根、冠及根冠间消长关系的影响,以期对调亏灌溉诱导作物补偿的机制进行初步探索。
1 材料和方法
1.1 试验地概况及试验材料
试验地点设在河南农业大学三区防雨棚内(37.80°N,113.65°E)。该地区年平均气温14.3 ℃,年降雨量640 mm,属于暖温带大陆性气候。供试玉米材料为父、母本自交系及其杂交1代(F1),其中父本(Male,M)为豫M808,属于兰卡系;母本(Female,F)为豫M5-236,属于瑞德系,两亲本中母本抗旱性更强。F1杂种优势显著。
1.2 试验设计
试验于2017年5-6月进行,采用桶栽种植模式(桶底直径15.4 cm,桶面直径19.5 cm,高18 cm)。供试土壤取自河南农业大学科教园区耕层土(全氮含量0.72 g/kg,有效磷含量9.8 mg/kg,速效钾含量124 mg/kg,有机质含量9.59 g/kg,碱解氮含量17 mg/kg),经过筛、晾晒、拌肥(复合肥)后,按容重1.2 g/cm3计算,每桶精确装干土3.7 kg,施肥量折合每桶3.8 g。播种前对桶内土壤均匀浇水,自然晾至田间持水量(Field capacity,FC)95%左右时播种,每桶播3粒种子,自播种之日起按80%FC供水。每个试验材料设计2个处理:连续干旱复水组(T)自植株3叶全展后,进行连续3次干旱复水控制,自然干旱至35%~45%FC(约3 d)后再复水至80%FC并持续复水3 d;对照组(CK)土壤在试验期内保持在65%~80%FC。控水期间采用称质量法补水,于每天17:00左右用精度为1.0 g的电子秤称每桶质量,根据水分减少量灌水至事先插入土中的PVC管中(PVC管埋入土中的部分钻有小孔,管外包裹细纱网以防小孔被土堵塞),避免破坏土壤表层结构、避免土壤板结。试验共种植118桶玉米,其中M、F、F1分别有40,39,39桶,每个材料设对照18桶,其余为处理。整个试验过程中每个材料处理间除水分存在显著差异外,其余管理条件保持一致。
1.3 测定指标及计算方法
1.3.1 田间持水量测定与日浇水量计算 取过筛后未拌肥的土壤,常温保存,烘干法测土壤田间持水量(FC),则日浇水量(g)=(80%FC时桶的总质量/kg-对应桶当日称质量/kg)×1 000。
1.3.2 植株冠层及根系性状测定 在第1次复水前(处理第3天)后(处理第6天)、第2次复水前(处理第9天)后(处理第12天)、第3次复水前(处理第15天)后(处理第18天)对各处理植株地上部分进行株高、叶面积的测量。株高为所有叶片自然伸展至最高处时的植株高度;叶面积采用估算法,测量每株全展绿色叶片的长度和最大宽度后,根据单叶面积=长×宽×0.75计算单叶面积。对每株单叶面积求和即为单株总叶面积。每个处理3个重复。
对最后一次取样后的根系,于烘干前用WinRhizo根系扫描系统测定根系形态指标。将根系(玉米根系较多,为避免根系在水槽内过多交叉,每个根样均被分成三部分进行扫描)摆入装有蒸馏水的无色透明水槽,用镊子调整根的位置尽量避免交叉重叠,使用根系扫描仪(型号为EPSON 1680)及其配套的根系分析软件扫描获取根系图像。用该软件分析根系各参数,根长、根体积、根系比表面积为上述三部分分析结果的总和,根系平均直径为上述三部分分析结果的平均值。每个处理3个重复。
1.3.3 植株生物量测定及根冠比的计算 在每次复水前后取玉米整株样,将根部洗净,与地上部分分离,分别于烘箱内105 ℃杀青1 h后80 ℃烘干至恒质量,分别称质量作为地下、地上生物量,并计算总生物量和根冠比。总生物量=地上生物量+地下生物量,根冠比=地下生物量/地上生物量。
1.3.4 生长恢复能力计算 生长恢复能力=ΔT/ΔCK[11];处理组植株:ΔT=复水后生物量-复水前生物量;对照组植株:ΔCK=复水后生物量-复水前生物量。
1.4 数据处理与统计分析
用Excel 2007对试验数据进行整理计算,用SAS 8.0中的t测验对数据进行方差分析(P≤0.05),用SigmaPlot 12.5作图。
2 结果与分析
2.1 连续干旱复水对玉米苗期总生物量的补偿效应
由图1可得,第1次干旱胁迫显著抑制了F材料总生物量的积累(P<0.05),对M材料无显著影响,但促进了F1材料总生物量的积累,处理T较对照略有增加;复水后,3个材料处理较复水前均有增加,但F材料并未增加至对照水平。第2次干旱胁迫时,F、M材料的生物量不减反增,F1材料却较对照略减;复水后,仅F材料的生物量出现处理T高于对照的现象,M、F1材料生物量分别较对照略减。第3次干旱胁迫下,M、F1材料的生物量较对照分别显著降低了22.7%,23.7%,而F材料的生物量较对照显著增加14.9%;第3次复水后,M材料的生物量较对照降低19.9%,而F、F1材料的生物量均值均高于对照,分别较对照增加了1.2%,1.4%。说明连续干旱复水显著抑制了M材料总生物量的积累,对F、F1材料生物量的补偿效果并不显著。总生物量仅是植株生长积累的总和,对物质分配及根冠补偿能力做进一步的分析才能更详细地了解植株在连续干旱复水这一过程中根、冠及根冠间发生的变化。

数据标以不同字母表示处理间差异显著。图2-5,7-8同。Values followed by different letter are significantly different at 0.05 probability level.The same as Fig.2-5,7-8.
2.2 连续干旱复水下玉米苗期根冠补偿效应的发生规律
经历干旱胁迫及复水后的植株会存在短暂的快速生长,但受基因控制,不同植株在复水后光合产物在根冠之间的分配比例会发生变化,这种变化可反映植物体内物质合成与分配是否受干旱胁迫及复水的影响以及植物在水分多变环境下的适应方式。
2.2.1 冠层生长补偿效应 植株的株高和叶面积是反映其生长状况的重要表型参数。由图2,3可知,株高与叶面积在连续干旱复水下的增减变化几乎是同步的,就F材料来说,第1次干旱复水过程中株高与叶面积的变化呈现出干旱下低于对照、复水后超过同时期对照的情况,但经受再次干旱胁迫后植株的株高在复水后并未恢复至对照水平,叶面积虽在第2次复水后略高于对照,但第3次复水后株高和叶面积分别显著低于对照18.1%,27.8%;M材料在第1次干旱复水过程中与F材料有相同的规律,从第2次复水开始,株高与叶面积在复水后的恢复程度没能再达到对照水平,最终导致第3次复水后分别显著低于对照10.9%,28.4%;干旱胁迫及复水对F1材料株高和叶面积的影响有延迟效应,第1次干旱胁迫并未显著影响F1材料株高及叶面积的增长,抑制作用出现在第2次干旱胁迫下,复水的促进作用也开始于第2次复水后,这种延迟效应,揭示了F1材料对环境的适应过程,其适应的结果是连续干旱复水后株高与叶面积分别较对照提高2.1%,13.6%,表现出超补偿效应。

图2 连续干旱复水过程中玉米株高的变化Fig.2 Variation of plant height during successive drought and re-watering in hybrid maize and its male and female parents

图3 连续干旱复水过程中玉米单株叶面积的变化Fig.3 Variation of leaf area during successive drought and re-watering in hybrid maize and its male and female parents
叶面积与株高在干旱复水下的生长变化也决定了地上生物量的积累。由图4可见,复水对F材料地上生物量积累的激发作用在第2次复水后较为突出,显著高于对照42.0%,呈现出超补偿效应,除此之外的2次复水后均表现为处理T略低于对照;M材料在连续干旱复水过程中地上生物量始终低于对照,最终的生物量较对照显著降低30.1%;较为特殊的是F1材料,连续干旱复水对其地上生物量积累呈现出促进作用,从第1次干旱胁迫与复水开始,地上生物量就高于对照,之后每个取样期处理T与对照均无显著差异,在第3次复水后,地上生物量略高于对照4.7%,有超补偿效应。F1材料的这种特殊性,一方面佐证了其超亲优势,一方面也体现了F1材料对连续干旱复水具有极强的适应及调节能力。

图4 连续干旱复水过程中玉米地上生物量的变化Fig.4 Variation of aboveground biomass during successive drought and re-watering in hybrid maize and its male and female parents
2.2.2 根系生长补偿效应 干旱复水对地上部生长的影响,其中很大程度上取决于根系的生长状况。由图5可知,在连续干旱复水下3个材料根系生长呈现出完全不同的响应规律。F材料在前2次干旱复水过程中根系生物量在逐步增加但始终低于对照,第3次干旱复水后却略高于对照3.9%,表现出超补偿效应;与F相反,M材料在前2次干旱复水中根系生物量受到干旱胁迫刺激和复水动力而得以快速增加,出现高于对照的情况,但第3次干旱复水后显著低于对照7.0%;F1材料根系生物量仅在第1次复水后略高于对照,再次受到干旱胁迫与复水时始终低于对照,第3次干旱复水后根系生物量略低于对照3.4%。

图5 连续干旱复水过程中玉米地下生物量的变化Fig.5 Variation of underground biomass during successive drought and re-watering in hybrid maize and its male and female parents
对第3次干旱复水后的根系进行分析可得图6,7,经历连续3次干旱复水后,对照组根系的上下层分布比较均匀,而处理组根系分布表现为下层多于上层(图6),且F材料根系的总根长、根系比表面积、根体积分别显著减少了15.9%,8.1%,31.0%,根系平均直径显著增加了10.0%。由图5可知,F材料地下生物量高于对照3.9%,可见其生物量高于对照的主要原因是原有根系增粗而不是毛细根的增加。M材料根系的总根长、根系比表面积、根体积分别增加了14.1%,3.4%,12.0%,根系平均直径显著降低了8.0%,且第3次复水后比复水前根系生物量增加了11.9%,达到了对照的93.0%,由根系生长特性分析可得,增加的这部分生物量主要依赖于老根的伸长和增粗。F1材料根系总根长、根系比表面积、根体积、根系平均直径分别较对照增加了6.3%,8.8%,27.1%,4.4%,具有显著超补偿效应,F1材料地下生物量在第3次复水后高于对照3.4%,较复水前增加了14.5%,可见,F1材料根系对连续干旱复水的响应主要体现在毛细根增加以及老根的伸长。

图6 第3次复水后玉米植株根系扫描图Fig.6 Scanning graphic of root after three times drought and re-watering
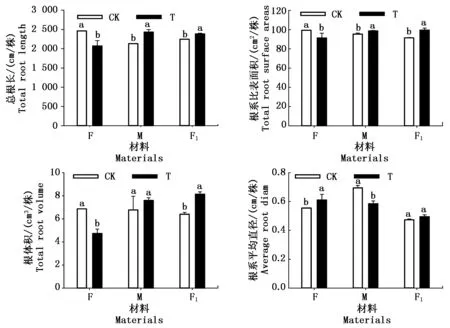
图7 三次干旱复水后玉米植株根系的形态特性Fig.7 Morphological characteristics of roots after three times drought and re-watering in hybrid maize and its male and female parents
2.2.3 根冠比及根冠生长恢复能力的变化 根冠比是揭示植株根冠之间物质分配消长关系的重要依据,由图8可得,就整个试验期而言,正常浇水条件下,玉米植株的根冠比随生育进程推进最终呈减小状态,以此来保证地上干物质的积累。但经过3轮干旱复水处理,第18天时M材料的根冠比表现为处理显著高于对照,提高了33.2%,说明M材料根系对干旱胁迫很敏感且干旱胁迫显著促进其根系生长;F材料在干旱复水条件下,地上干物质增加率显著高于根系增加率,因此自第6天始,根冠比显著低于对照直至第12天;处理第15天时与对照的生长特征基本同步,且第3次干旱复水后根冠比较对照高了5.8%,说明在干旱胁迫下,F植株的物质分配量为地上>根系,这与冠层生长(叶面积和株高)的变化一致;F1材料结合了2个亲本的优良性状,在整个干旱复水过程中,物质分配倾向为先根系后冠层,第3次干旱复水后根冠比较对照降低7.3%,前期根系的良好发育为后期冠层的超补偿奠定了基础。

图8 连续干旱复水过程中玉米根冠比的变化Fig.8 Change of root-shoot ratio during successive drought and re-watering in hybrid maize and its male and female parents
为更好地评价植株的生长恢复能力,分别计算了每次复水后地上、地下生物量的ΔT/ΔCK值,以1.0为标线,ΔT/ΔCK≥1.0时为优;ΔT/ΔCK<1.0时为劣。对地上生长而言,图9-A显示,F材料在3次复水后生长恢复能力递减,前2次复水后虽均达到了超补偿水平,但第3次干旱复水后地上生长恢复能力降至标线以下接近于0;M材料前2次复水后生长恢复能力均处于标线以上且基本无差异,但第3次复水后生长恢复能力下降至标线以下,说明在本试验设计水平下,连续3次干旱复水对亲本F、M冠层的物质积累是不利的;F1材料冠层的生长恢复能力随着复水次数的增加呈递增趋势,且每次都处于标线以上,即每次复水后地上物质积累都产生了超补偿作用且干旱复水次数的增加能增大其超补偿程度。结合地下根系在复水后的生长恢复能力(图9-B)可以发现,两亲本F、M根系的补偿生长能力显著强于F1。两亲本F、M根系的生长恢复能力随胁迫次数增加呈递减趋势,F1材料根系的生长恢复能力在第2次复水后得到显著提高,但第3次复水后迅速降低,这可能是两亲本在第3次复水后物质分配趋向于根系,F1更趋向于地上部分的原因。

A.冠层; B.根系。A.Shoots; B.Roots.
3 结论与讨论
本试验结果表明,连续干旱复水对父母本自交系及其F1玉米苗期生物量积累有不同影响。F1和耐旱型母本在试验结束时生物量虽略高于对照但并无显著差异,其中F1增幅略高于母本,且F1净生物量显著高于母本,说明F1的整体抗旱性强于母本。这与慕自新等[12-13]的研究结果相似,其在对一组玉米遗传材料的系统研究中发现,杂交种在水分利用上具有开源节流的作用,即在水分亏缺条件下,根系吸水能力增强,但相应的气孔导度降低,蒸腾作用减弱,减小了蒸腾的冗余部分,使水分得到有效利用,物质合成及积累呈现较强优势,表现出显著补偿效应;水分胁迫解除后,这一优势得到了更充分的发挥,体现了F1较强的超亲优势和适应性。
植物对变水环境的适应性主要是通过调整根冠关系来实现的,而根冠关系是由遗传背景和环境因子共同决定的。大多研究表明,经过水分胁迫及复水的植物根冠比会增大[14-15],但本研究结果显示,F1材料的根冠比在经历连续干旱复水后反而低于对照。在受到干旱胁迫时,植物生物量倾向于分配到根系,以增加吸水,满足地上部分的蒸腾需求,也使根系保持一定的活力[16]。复水后,由于植株水分需求被满足,促使地上部分进行正常的光合作用,同时光合产物也进行了再分配,再分配部分干物质补偿了植株在干旱胁迫下受到的伤害[17]。但生产单位根系所需光合同化产物约是形成单位地上部干质量的2倍[18],因此,F1材料在复水后物质分配趋向于冠层,降低了生长冗余,根系结构更适宜,其吸收水分、养分的功能和叶冠的光合作用配合最恰当,提高了光合产物的利用率[19]。两亲本中的母本在前2次干旱复水过程中呈现与F1相似的物质分配规律,但第3 次干旱复水时根冠比高于对照,可能是前2次干旱复水并未解除前期干旱胁迫对母本材料冠层的伤害,累积的伤害在第3次干旱复水后显现出来。而连续干旱复水下的父本材料根冠比始终高于对照,原因在于复水后物质分配趋向于强根弱冠。
根系在根冠关系中扮演着更为重要的角色,根系对土壤水分的吸收和利用也在一定程度上决定了冠层的生长情况。王空军等[20]、孙庆泉等[21]对我国玉米品种更替过程中根系特征的演变进行了系统研究,结果发现,现代玉米品种普遍呈现“横向紧缩、纵向延伸”的根系特征,这种特征使得土壤上层的根量相对减少,下层根系数量多,能够在上层土壤干旱时从深层吸水。李国领等[22]在研究中也发现,具有更高的根系干物质量可能是小麦获得高产及优良抗旱特性的原因。而在桶栽玉米试验中,由于根系生长空间有限,根系这一特性使得植株在试验所设水分胁迫条件下也能吸收水分保证生长。本试验所得根系扫描图可发现,在正常水分条件下,上下层根系分布较为均匀,而经历连续干旱复水的植株根系下层根量显著多于上层,且F1材料冠层生长恢复能力随干旱胁迫频次增加而增强,这在于干湿交替后,一方面根尖恢复生长,长出大量新根,并产生根毛,根系纵向生长加强而深扎,有利于吸水;另一方面,由于大量新根的出现和胁迫激素ABA的作用,干旱复水后根系的水分传导增大。此外,根从湿润土壤吸水可通过水分倒流维持干土中根系的功能[23],避免干土中根系的衰老,为复水后这部分根的超补偿吸水奠定了基础。这些保证了根和叶延伸生长区膨压在一定程度上的维持,也就保证了复水后各种生理功能的快速恢复和地上部的补偿生长。
综上所述,连续干旱复水下玉米杂交种具有显著的超亲优势,根系特性上遗传并融合了父母本各自的特征,下层根系更为丰富;根冠关系上继承并优化了母本的根冠物质分配,冠层补偿逐渐增加,根系补偿弱化,保障了地上部在多变水分环境下的生长。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
草业科学(2022年2期)2022-03-21
农业科技通讯(2022年2期)2022-02-28
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
新疆农业科学(2020年1期)2020-02-14
中国科技纵横(2018年14期)2018-08-30
现代农业科技(2017年16期)2017-09-22
