
盐胁迫对寒地粳稻籽粒淀粉形成积累及产量的影响
2019-03-08 02:00胡博文谷娇娇沙汉景张君颜黄书勤赵宏伟
华北农学报 2019年1期
胡博文,谷娇娇,贾 琰,沙汉景,张君颜,黄书勤,赵宏伟
(东北农业大学 水稻研究所,黑龙江 哈尔滨 150030)
黑龙江省是寒地粳稻的重要栽植区域,其种植总面积及产量稳步持续增长[1]。盐胁迫对水稻有诸多影响,其中包括:造成发育延迟、抑制水稻组织与器官生长分化,最终影响产量及品质[2-4]。淀粉是水稻籽粒中的重要组成成分,其比例大多为80%~90%,因此,淀粉的形成积聚过程即为水稻籽粒的充实过程[5],而该过程受盐胁迫的影响较大。
籽粒中的淀粉是由源器官所产生的光合产物,以蔗糖的方式运输到籽粒后在多个酶的共同作用下转化而成[6-7]。这一过程中,ADPG焦磷酸化酶、可溶性淀粉合成酶(SSS)以及淀粉分支酶(Q酶)发挥主要调控作用[8-9]。ADPG焦磷酸化酶的功能是产生直接前体ADPG,催化葡萄糖-1-磷酸与ATP的反应,该酶是淀粉合成过程中的限速酶[10]。在SSS的催化作用下,生成的ADPG与淀粉引物(葡聚糖)发生相互作用从而使葡萄糖分子转运至淀粉引物,致使淀粉链的延伸[11]。Q酶决定淀粉组分与结构,是支链淀粉合成的关键酶[12]。
当前,国内外已有许多学者对盐胁迫下水稻的响应机制进行了深入研究。陈海燕等[13]研究表明,在盐胁迫下水稻根系中超氧化物歧化酶(SOD)和抗坏血酸过氧化物酶(APX)活性无明显变化,而耐盐品种的过氧化氢酶(CAT)和过氧化物酶(POD)活性高于盐敏感品种。晏斌等[14]发现,在86 mmol/L的NaCl培养液中,适当增加外界 Ca2+浓度,会使水稻体内Na+浓度下降,并保证水稻植株有较高相对生长率。Wutipraditkul等[15]研究发现,施用外源物质脯氨酸可增强水稻的耐盐性。前人多从抗氧化机制、渗透调节以及离子分布等角度解析水稻对盐胁迫的响应机制,但盐胁迫对灌浆结实期寒地粳稻籽粒淀粉合成关键酶活性、淀粉含量及最终产量形成的影响的相关研究仍缺乏系统性报道。
本试验采用寒地粳稻品种龙稻5(耐盐)、牡丹江30(盐敏感)为试验材料,研究不同浓度盐胁迫下寒地粳稻籽粒淀粉合成关键酶活性变化规律及其与淀粉含量间关系,揭示寒地粳稻籽粒淀粉合成代谢对盐胁迫的响应机制,明确盐胁迫对寒地粳稻产量及产量构成因素的影响,以期丰富寒地粳稻耐盐研究的生理基础,并为水稻选择适宜的盐碱地种植和盐渍土水稻栽培提供理论依据。
1 材料和方法
1.1 试验材料
经东北农业大学水稻研究所前人筛选所得的耐盐性不同的2个寒地粳稻品种:牡丹江30(盐敏感)、龙稻5(耐盐)。
1.2 试验设计
试验在东北农业大学盆栽场进行。于2017年4月20日播种,5月30日移栽。本试验采用完全随机试验设计,选择无渗漏的塑料盆体,土壤风干后过筛,每盆装10 kg土壤。施盐量设置5个梯度,以施入土壤NaCl占土壤总量的百分比表示,分别为:0,0.075%,0.150%,0.225%,0.300% 。其中,牡丹江30各处理依次记为M1(对照)、M2、M3、M4、M5,龙稻5则记为L1(对照)、L2、L3、L4、L5。共计10个处理。于返青后开始盐处理,分5次处理完毕。盆栽密度设置为4穴/盆,每穴3株,每个处理80盆。田间管理同一般生产田。
1.3 试验方法
1.3.1 籽粒淀粉合成代谢关键酶活性及淀粉的取样方法 记录各处理到达齐穗日期,并于齐穗期标记每处理生长整齐的植株120穴。在齐穗后的7,14,21,28,35 d的9:30-11:00(晴天)选取每处理所标记植株6穴,其中,3穴取稻穗中部籽粒用液氮冷冻处理并移至-80 ℃冰柜,用于测定籽粒的ADPG焦磷酸化酶、SSS和Q酶活性。剩余3穴籽粒用于淀粉含量的测定。
1.3.2 测定方法
1.3.2.1 籽粒淀粉合成代谢关键酶的测定方法 参照李太贵等[16]测定Q酶活性的方法。以吸光值下降一个百分点为一个酶活性单位U。
参照Doehlert等[17]、程方民等[18]方法,采用相应试剂盒测定ADPG焦磷酸化酶、SSS活性。酶活性测定重复3次。
1.3.2.2 籽粒淀粉及其组分含量测定 称取各处理籽粒粉碎样品0.1 g用于淀粉含量的测定,3次重复。采用双波长法测定籽粒淀粉含量[19]。以每1 000 g籽粒中淀粉量的克数表示,即g/kg。
1.3.2.3 产量及产量构成因素测定 在成熟期每处理选取9穴测定有效穗数、穗粒数、千粒质量及结实率并计算理论产量。
1.4 数据分析
采用Microsoft Excel 2010和SPSS 19.0统计分析数据。
2 结果与分析
2.1 盐胁迫下寒地粳稻籽粒淀粉组分的动态变化
2.1.1 直链淀粉积累动态 由图1可知,寒地粳稻籽粒直链淀粉含量随齐穗后天数的递增呈持续增长状态,直链淀粉含量的增长趋势由快速增长趋于平缓增长。同对照相比(M1、L1),盐处理下籽粒直链淀粉含量处于较高水平。牡丹江30的M1及M5表现为:齐穗后7 d差异不显著,其余天数均表现为差异显著;龙稻5的L1及L5表现为:齐穗后7,14,21 d差异不显著,其余天数差异显著。
在齐穗后的7~14 d这一快速增长阶段,盐敏感品种牡丹江30各处理增长速率为 M4>M5>M1>M2>M3,耐盐品种龙稻5为L3>L5>L1>L4>L2,当土壤含盐量达到0.300%时,2个品种的直链淀粉增长速率均高于对照水平。2个寒地粳稻品种在高盐胁迫下的反应不同,具体表现为增长速率M4>L4、M5>L5。
齐穗后21~35 d籽粒直链淀粉含量增长趋于平缓。牡丹江30籽粒直链淀粉含量在齐穗后35 d表现为M4>M5>M3>M2>M1;龙稻5为L5>L4>L3>L2>L1。

图1 盐胁迫下籽粒直链淀粉含量动态变化Fig.1 Dynamic changes of amylose content in kernels under salt stress
2.1.2 支链淀粉积累动态 由图2可知,寒地粳稻籽粒支链淀粉的积累规律为持续增长趋势。盐胁迫降低寒地粳稻籽粒支链淀粉含量,具体表现是土壤含盐量越高,籽粒支链淀粉含量越低。牡丹江30的M1及M5表现为:齐穗后7 d差异不显著,其余天数均表现为差异显著;龙稻5的L1及L5表现为:除齐穗后7,21 d差异不显著外,其余天数均差异显著。
可以发现,在齐穗后的7~21 d这一阶段,2个寒地粳稻品种的各处理在积累支链淀粉含量的速率上表现不尽相同,其中M1、M2、L1、L2、L3处理表现为先快速增长后缓慢增长,其余处理则是先缓慢增长后快速增长。可见,当土壤含盐量达到0.150%时,牡丹江30受影响较大,而龙稻5则是当土壤含量达到0.225%时才受较大影响。
当灌浆过程进行到齐穗后21 d后,各处理增长速率趋于平缓。对照及低浓度(0.075%)盐处理下寒地粳稻籽粒支链淀粉含量一直保持相对较高水平,且差异较小。

图2 盐胁迫下籽粒支链淀粉含量动态变化Fig.2 Dynamic changes of amylopectin content in kernels under salt stress
2.2 盐胁迫下籽粒淀粉及其组分含量分析
寒地粳稻籽粒淀粉及其组分含量(齐穗后35 d)分析如表1所示。M4处理和L5处理下寒地粳稻籽粒直链淀粉含量最高,且均显著高于对照水平。 2个供试品种在不同浓度盐胁迫下的应激能力不同,牡丹江30各处理籽粒直链淀粉含量较对照的增幅较大,为2.43%~9.72%,龙稻5增幅较小,为1.42%~9.00%。
与对照水平相比,盐胁迫下寒地粳稻籽粒支链淀粉含量下降,除M2、L2处理与对照差异不显著外,其余各处理均显著低于对照水平。总淀粉含量变化趋势同支链淀粉。盐胁迫下牡丹江30籽粒支链淀粉、总淀粉含量较对照的下降幅度分别为1.09%~18.62%和0.25%~12.09%,龙稻5分别为0.58%~15.46%和0.12%~9.9%。可以发现,大量的NaCl堆积严重影响籽粒淀粉形成,且品种对盐胁迫的反应有所不同,耐盐品种龙稻5的表现好于盐敏感品种牡丹江30。从M1与M2、L1与L2可以看出,总淀粉含量以及淀粉组分含量都表现为差异不显著,这充分说明寒地粳稻可抵御来自低浓度盐胁迫所带来的毒害。
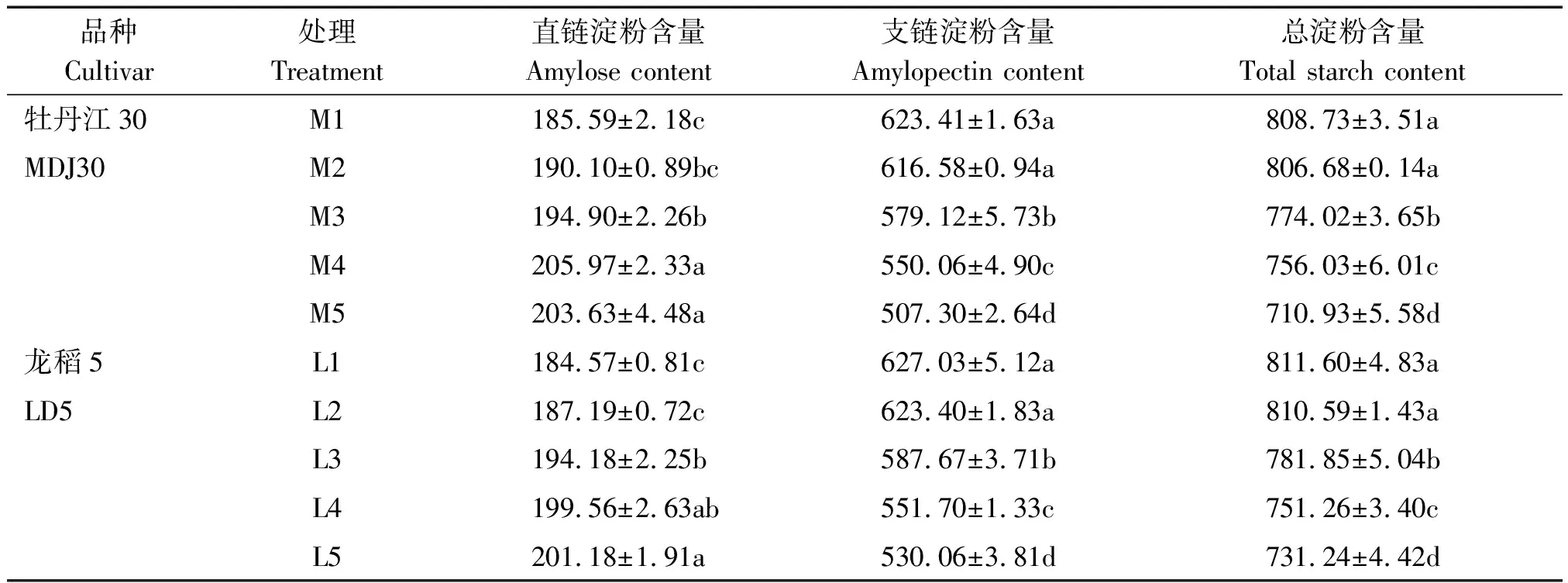
表1 盐胁迫下籽粒淀粉及其组分含量Tab.1 Starch component contents in kernels under salt stress g/kg
注:表中不同的小写字母表示5%水平的差异显著。表3同。
Note: Values followed by different small letters are significantly different at the 5% probability levels. The same as Tab.3.
2.3 盐胁迫对淀粉合成关键酶活性的影响
2.3.1 淀粉分支酶(Q酶)的动态变化 如图3可知,随着齐穗后天数的增加,寒地粳稻籽粒Q酶活性(以鲜质量计)体现为单峰曲线变化趋势。但各处理Q酶活性抵达峰值的日期不同,于齐穗后21 d抵达峰值的处理有:M1、M2、L1、L2、L3,其中M1同M2差异不显著,L1同L2、L3差异显著;其余处理于齐穗后28 d达到峰值,其中M3、M4、M5差异不显著,L4、L5差异不显著。2个品种籽粒Q酶活性对盐胁迫的响应不同,盐敏感品种牡丹江30在土壤含盐量达到0.150%时,Q酶活性严重受抑,出现峰值后移现象,而耐盐品种龙稻5则是在土壤含盐量达到0.225%才出现以上现象。同时,随着土壤含盐量的增高,酶活性峰值下降明显,其具体表现为M1>M2>M4>M3>M5、L1>L2>L3>L4>L5。

图3 盐胁迫下寒地粳稻籽粒Q酶活性的变化规律Fig.3 Changes of Q activity in kernels of Japonica rice in cold-region under salt stress
2.3.2 可溶性淀粉合成酶(SSS)的动态变化 如图4可知,寒地粳稻籽粒SSS活性(以鲜质量计)随齐穗后天数的增加呈单峰曲线变化,但各处理峰值出现的日期不尽相同,具体表现为:M1、M3、L1、L2、L3处理于齐穗后14 d达峰值,其中,M1同M3差异不显著,L1同L2、L3差异不显著;其余处理均在齐穗后21 d达峰值,且在齐穗后21 d达峰值的品种内各处理均表现为差异不显著。通过对比2个品种可以看出,当土壤含盐量达到0.075%时,牡丹江30受影响较大,峰值日期推迟(除M3处理);龙稻5表现较好,当土壤含盐量达到0.225%时,才出现峰值后移现象,其峰值具体表现为M1>M4>M2>M5>M3、L2>L1>L4>L5>L3。
2.3.3 ADPG焦磷酸化酶的动态变化 由图5可知,寒地粳稻籽粒ADPG焦磷酸化酶活性(以鲜质量计)随生育进程的推进表现为单峰曲线,但各处理到达峰值日期存在差别。M1、M2、M3、L1、L2、L4在齐穗后14 d达峰值,其中M1同M2、M3差异显著,L1同L2、L4差异显著;其余处理在齐穗后21 d达峰值,其中M4、M5差异显著,L3、L5差异不显著。盐胁迫下籽粒ADPG焦磷酸化酶活性峰值变化趋势具体表现为M1>M2>M3>M4>M5、L1>L2>L3>L5>L4。且耐盐品种龙稻5各处理峰值酶活性高于盐敏感品种牡丹江30各处理(除M4、L4外)。
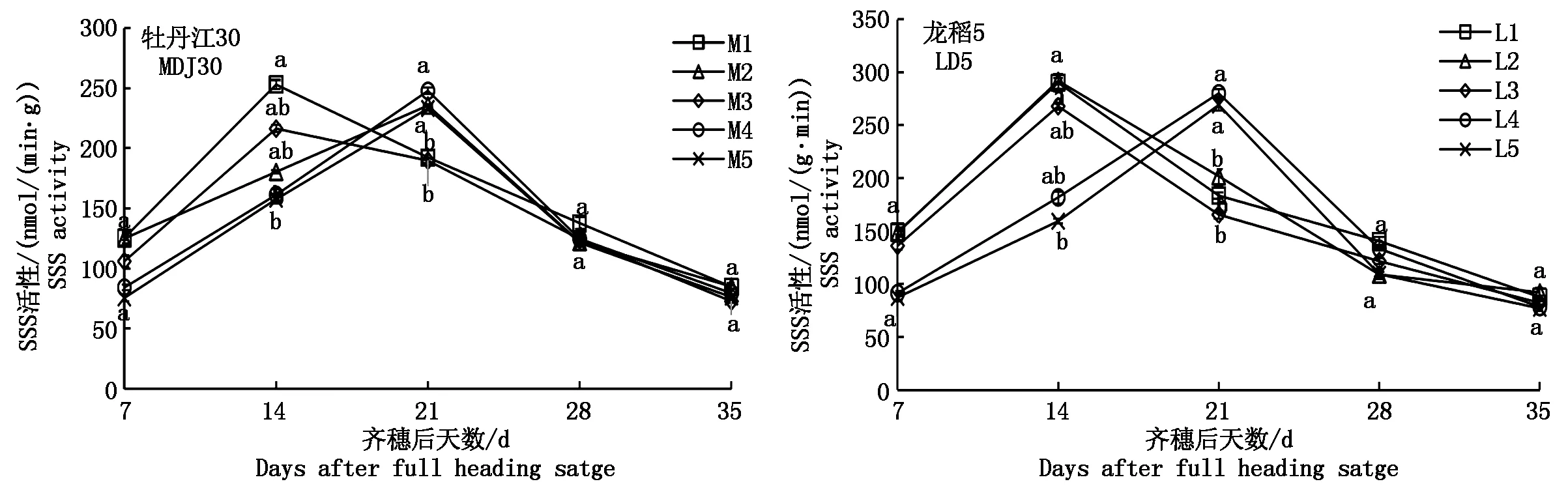
图4 盐胁迫下寒地粳稻籽粒SSS活性的变化规律Fig.4 Changes of SSS activity in kernels of Japonica rice in cold-region under salt stress

图5 盐胁迫下寒地粳稻籽粒ADPG焦磷酸化酶活性的变化规律Fig.5 Changes of ADPG-PPase activity in kernels of Japonica rice in cold-region under salt stress
2.4 淀粉合成关键酶活性与淀粉及淀粉组分之间的相关分析
如表2所示,寒地粳稻籽粒支链淀粉含量以及总淀粉含量与齐穗后14,21,28,35 d的Q酶活性呈显著或极显著正相关,与齐穗后14,21,35 d的SSS活性呈显著或极显著正相关,可以发现,籽粒Q酶与SSS共同完成支链淀粉的合成,通过影响支链的淀粉含量最终决定总淀粉含量。籽粒总淀粉含量与ADPG焦磷酸化酶在齐穗后7,35 d呈极显著正相关,这说明ADPG焦磷酸化酶主要作用于籽粒灌浆过程的前期与后期。

表2 盐胁迫下寒地粳稻籽粒 ADPG 焦磷酸化酶、SSS、Q 酶活性与淀粉含量之间的关系Tab.2 Relationship of ADPG-PPase, SSS, Q enzyme activities, and starch contents in kernels of Japonica rice in cold-region under salt stress
注*、**分别表示在0.05和0.01水平上显著相关。
Note*,**indicate significant correlation at the 0.05 and 0.01 levels,respectively.
2.5 盐胁迫对寒地粳稻产量及产量构成因素的影响
由表3可见,盐胁迫下2个品种的各产量构成因素及理论产量较对照(M1、L1)均有不同程度下降。牡丹江30各处理有效穗数的降幅为0~27.08%,龙稻5为4.65%~41.86%,具体表现为:M1同M2、M3差异不显著,但显著高于M4、M5;L1同L2差异不显著,但显著高于L3、L4、L5。牡丹江30穗粒数的降幅为7.32%~39.46%,龙稻5为4.69%~25.51%。其中,M1的穗粒数显著高于各处理;L1与L2、L3处理差异不显著,但显著高于其余处理。
牡丹江30的结实率,各处理较对照下降0.02~14.33百分点,而龙稻5则为1.20~11.45百分点。同时,牡丹江30的千粒质量、理论产量降幅依次下降为:7.97%~22.98%,14.41%~71.30%;龙稻5为1.24%~15.08%,10.92%~67.49%。2个品种的处理间表现相同,即当土壤含盐量达到0.150%时,结实率及理论产量显著下降;当土壤含盐量达到0.225%时,千粒质量显著下降。
通过分析可知,M1和M2、L1和L2处理各指标均表现为差异不显著(除牡丹江30的穗粒数),说明水稻可承受较低浓度的盐胁迫。同时可以发现,牡丹江30的穗粒数及结实率在处理间差异较大,而龙稻5则是有效穗数及结实率差异较大,说明耐盐性不同的品种在盐胁迫下产量构成因素的变化存在差异。通过对比2个品种,除有效穗数外,牡丹江30的各产量构成因素降幅均高于龙稻5,有效穗数降幅较小的原因可能是牡丹江30具有相对较好的分蘖能力。
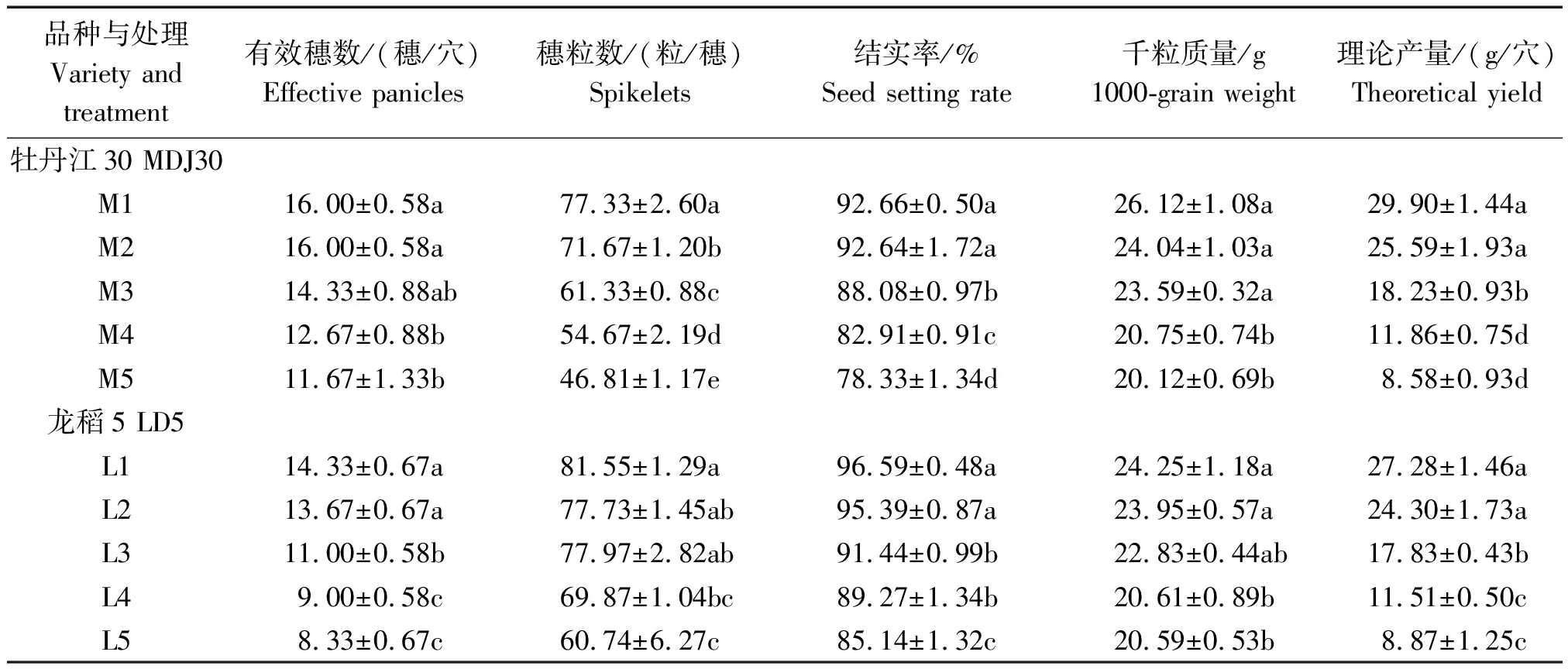
表3 盐胁迫对寒地粳稻产量构成因素的影响Tab.3 Effects of salt stress on yield components of Japonica rice in cold-region
3 结论与讨论
在一系列淀粉合成相关酶的调控作用下,籽粒中淀粉开始形成并逐步积累,这一动态变化即为水稻籽粒充实的过程。已有大量研究指出,ADPG焦磷酸化酶、SSS、Q酶在高等植物的光合及非光合器官中都发挥着至关重要的调控功能,同时,由NaCl引起的不良影响并不会随NaCl的消失而减弱[20]。本研究发现,随土壤含盐量的提升,峰值酶活性波动较大。这可能是在高浓度盐胁迫下(0.150%~0.300%),植物体内由于累积了大量Na+使得原有的离子平衡被破坏,在一定程度上损伤了植物的基础蛋白质结构,从而使酶活性大幅下降[21]。通过淀粉合成关键酶动态变化可看出,盐胁迫延迟酶活性到达峰值的日期,分析原因可能是作物本身需要时间来适应盐胁迫。有研究指出耐盐品种较盐敏感品种更能有效地控制外界Na+向地上部的转运,可在一定范围避免地上部遭受盐害[22-23]。通过导致淀粉合成关键酶活性峰值推迟的土壤含盐量来看,耐盐性品种龙稻5更能有效避免Na+所带来毒害,在盐胁迫下籽粒仍能保持相对较高的淀粉合成关键酶活性。
稻米品质取决于淀粉的组分与含量。步金宝[24]研究发现,盐处理显著抑制粳稻籽粒淀粉的最终形成,且淀粉组分积累速率受到影响。本研究发现,随着盐浓度的升高,2个供试品种籽粒的总淀粉及支链淀粉含量降幅逐渐增大,低浓度盐处理(M2与 L2)与对照差异不显著,而中高浓度盐处理导致总淀粉及支链淀粉含量下降明显,但具体降幅因品种的耐盐性而异。本研究同时还发现,盐胁迫可导致籽粒直链淀粉含量显著增加,直链淀粉含量的增幅随着盐浓度的升高而增大,这与罗成科等[25]的研究结果基本一致。分析原因可能是由于植株在遭受盐胁迫时可促进直链淀粉合成,从而改变直链淀粉与支链淀粉的比例,以此来抵御不良环境带来的影响,但这一现象是否为水稻的一种应激反应还有待进一步研究。
通过试验结果分析可知,籽粒支链淀粉含量与Q酶活性呈显著或极显著正相关(除齐穗后7 d),并与 SSS活性表现为灌浆中后期显著或极显著正相关,说明盐胁迫下寒地粳稻籽粒支链淀粉的合成与 SSS和Q酶关系密切,这在前人的研究中已有体现[26-27]。籽粒淀粉含量与灌浆前期、后期ADPG焦磷酸化酶活性呈极显著正相关,淀粉及其组分在灌浆前期都有一个快速增长的过程,说明 ADPG焦磷酸化酶的活性与灌浆速率密切相关,这和前人的研究结果基本一致[28-29],后期呈极显著正相关可能是由于当灌浆过程进行到后期,植株趋于成熟启动衰老机制,ADPG焦磷酸化酶通过抑制淀粉合成速率来减缓代谢。
产量形成依靠籽粒干物质的积累,又取决于籽粒淀粉合成积累的多寡[30]。本研究发现,2个供试品种的理论产量在土壤含盐量为0.075%时与对照差异不明显,当土壤含盐量达到0.150%时抑制效果明显。关于盐胁迫对水稻产量构成因素影响的说法不一:杨福等[31]研究得出盐碱胁迫下产量的下降是由穗粒数减少及千粒质量减轻所引起;余为仆[32]通过对产量构成因素的分析发现,盐胁迫导致的降产在多方面因素的共同作用下完成。本研究结果表明,产量构成因素各指标随土壤含盐量的提高而下降,但低浓度的盐处理(M2、L2)与对照差异不显著(除牡丹江30的穗粒数),当土壤含盐量超过0.075%时,各产量构成因素受影响显著,可以发现,结实率的变化最为突出。但寒地粳稻耐盐临界值是否为0.075%还需进一步探讨。此外,本研究结果为盆栽试验所得,可对大田生产有一定指导,但具体应用还有待进一步验证。
猜你喜欢
园林科技(2021年2期)2022-01-19
牡丹江医学院学报(2021年6期)2021-11-18
农业机械学报(2021年10期)2021-11-09
当代水产(2021年6期)2021-08-13
活力(2019年17期)2019-11-26
文萃报·周五版(2019年38期)2019-09-10
北京航空航天大学学报(2019年3期)2019-04-08
冰雪运动(2018年3期)2018-12-29
文苑(2018年21期)2018-11-09
食品工业科技(2014年13期)2014-12-16
