
番茄SlETR6基因的克隆及非生物胁迫下的表达分析
2019-03-08 02:00苏丽艳
华北农学报 2019年1期
苏丽艳
(西安文理学院 生物与环境工程学院,秦岭野生观赏植物研究中心, 陕西 西安 710065)
乙烯是一种小分子气态植物激素,对果实成熟、种子萌发、叶和花衰老等多种植物发育过程有重要调控作用[1-2]。此外,乙烯在植物响应生物和非生物胁迫过程中有重要的作用,当植物体受到冷害、干旱、损伤、低氧等生物及非生物逆境胁迫时,其体内乙烯的含量往往会增加[3]。乙烯信号通过相应的转导途径进行胁迫信号的传递,调控下游基因的表达进而使植物体在生理水平上发生一系列适应环境的改变。乙烯受体 (Ethylene receptor, ETR) 蛋白是乙烯的负调控因子,是乙烯信号转导途径上游重要组成元件之一,在缺乏乙烯时,受体会抑制乙烯诱导的相关基因表达;乙烯存在时,乙烯与受体蛋白结合使受体失活,从而激活下游的乙烯诱导相关基因[4-5]。
ETR家族包含多个同源基因,拟南芥中分离得到5个ETR受体基因,根据其结构域可分为两大亚家族,ETR1亚家族包括AtETR1和AtERS1,ETR2亚家族包括AtETR2、AtERS2、AtEIN3[6]。ETR1亚家族含有组蛋白激酶所需要的5个完整结构域,而ETR2亚家族基因仅含有部分组蛋白激酶所需要的结构域。番茄中分析筛选到6个乙烯受体,分别是:SlETR1、SlETR2、NR(Never-ripe)、SlETR4、SlETR5和SlETR6,它们在蛋白质水平上差异较大,在序列上表现出50%以下的同源性[7-10]。目前已经在凤梨、苹果、草莓、桃及芒果等多种植物中被分离出来[11-16]。
目前,对于乙烯受体ETR的研究多集中在其对植物生长发育的调控作用,如番茄乙烯受体SlETR1和SlETR2在乙烯信号转导中起负调控作用[17-18];NR突变体果实不能正常成熟[19];SlETR4的存在可促使番茄果实乙烯响应系统Ⅱ中乙烯的加速合成,促进果实早熟[20-21],果实成熟期时,SlETR5的表达明显上升,而SlETR6的功能可能与SlETR4相似。目前,对于ETR在胁迫过程中的功能只有少量报道,前期研究发现,AtEIN2突变后影响拟南芥植株的盐敏感度,证明AtEIN2参与盐胁迫信号的调控过程[22];可可CcEIN4抑制表达后可以提高植物对盐胁迫的抗性,表明乙烯受体CcEIN4亦参与调控植株的盐胁迫信号调控过程[23]。
番茄是研究呼吸跃变型果实发育特征的典型模式植物。SlETR6是番茄乙烯受体家族中发现较晚的一个成员,目前尚无SlETR6对干旱、高盐、温度胁迫响应的相关报道。本研究对番茄矮化品种Mic-Tom中SlETR6基因进行了克隆,并研究了SlETR6在不同植物组织及高盐、高温、低温、干旱等胁迫条件下的表达模式,旨在为进一步了解SlETR6在植物响应环境胁迫中的功能奠定理论基础。
1 材料和方法
1.1 试验材料及胁迫处理
Mic-Tom番茄种子为中科院遗传所馈赠。种植于西安文理学院植物生长繁育单元-西北文络型连栋温室中,番茄生长条件:白昼温度分别为光周期为16 h/8 h,昼夜温度为27 ℃/19 ℃, 湿度为60%~80% RH。取番茄植株的不同植物组织,包括根、茎、叶、花、种子和不同发育时期果实(小果/花后5 d、中果/花后10 d、大果/花后25 d、转色果/35 d、红果/花后45 d),取样后立即用液氮冻存并存于-80 ℃冰箱备用。
选取盆栽长势一致的Mic-Tom番茄35 d苗植株进行非生物胁迫处理,包括200 mmol/L NaCl溶液灌根、高温(40 ℃)、低温(4 ℃)、脱水即模拟干旱(将番茄苗洗净根部泥土置于滤纸上),以未做任何处理的植株为对照。每个处理3次重复,分别于处理后0,1,3,6,12,24 h收取叶片组织,液氮冻存后存于-80 ℃冰箱备用。
1.2 番茄总RNA提取及SlETR6基因的克隆
试验材料总RNA提取使用天根生化科技有限公司RNA提取试剂盒, 用1%琼脂糖凝胶电泳测定其浓度和完整性,利用DNase去除基因组DNA后,利用TaKaRa反转录试剂盒按照产品操作按说明书合成cDNA。
以番茄叶片cDNA为模板进行SlETR6基因ORF全长克隆,利用Primer 5.0设计引物SlETR6-F:5′-GCAATGATGAAGAAAGTA-3′,SlETR6-R:5′-GTC ATGGCATTCCTCTG-3′。采用TaKaRa 公司的PrimeSTAR®HS DNA Polymerase高保真酶进行扩增,反应体系为25 μL,反应程序是:94 ℃,5 min;94 ℃,30 s,58 ℃,30 s,72 ℃,90 s,34个循环; 72 ℃,10 min。PCR产物在1%琼脂糖凝胶电泳检测,将检测到的目的片段用琼脂糖凝胶纯化回收,将回收到的目的基因产物连接到pMD19-T载体后,转化大肠杆菌,筛选阳性克隆提取质粒后进行DNA测序检测。
1.3 番茄SlETR6基因的生物信息学分析
利用 DNAMAN 对SlETR6基因进行氨基酸序列比对;蛋白质分子质量和等电点预测利用在线软件 ProtParam(http://web. expasy.org/protparam/)计算完成;利用TargetP 1.1 Server、plantCARE 在线工具完成SLETR6蛋白的亚细胞定位预测、启动子顺式作用元件分析。利用 MEGA 5 软件构建SlETR6基因系统进化树。
1.4 SlETR6表达特性分析
对番茄不同植物组织及不同胁迫处理得到的材料提取RNA, 纯化后反转录为cDNA。在ABI Quant Studiotm6 Flex Real-time PCR 系统进行qPCR 分析。以cDNA为模板,以SlUBI(GenBank登录号:Q96483)为内参。用 Primer 5.0 设计引物,qSlETR6-F:5′-GAATGGCATCCACGGAG-3′,qSlETR6-R: 5′-CAATCTCCTCAGTGTCG-3′;SlUBI-F: 5′-TGT CCCTATCTACGAGGGTTAT-3′;SlUBI-R:5′-AGTTAA ATCACGACCAGCAAGAT-3′。主要步骤按荧光定量试剂盒说明书操作完成。反应总体系为20 μL,10 μL 荧光染料反应混合液,1 μL 模板cDNA (浓度 30 ng/μL),引物各1 μL (10 μmol/L),ddH2O 7 μL。反应程序为:95 ℃变性3 min;然后,95 ℃ 20 s,60 ℃退火30 s,72 ℃延伸30 s,共39次循环。qRT-PCR 反应于ABI Quant Studiotm6 Flex Real-Time PCR System上进行。相对表达量用2-ΔΔCT法分析[24]。
2 结果与分析
2.1 SlETR6基因的克隆和启动子序列分析
利用PCR方法克隆得到2 200 bp左右与预期大小一致的目标片段(图1)。测序结果表明,目标条带大小2 265 bp,与引物设计预测条带一致,编码754个氨基酸,其组成蛋白质分子质量为85.05 ku, 等电点为7.28。启动子序列分析表明,SlETR6含有TATA 框、CAAT 框等调控真核基因转录的基本元件外,还具有热胁迫、干旱胁迫、低氧胁迫、光响应、激素(乙烯、水杨酸、赤霉素)诱导响应、防御与胁迫响应元件等多种功能响应元件(表1)。
2.2 SlETR6同源序列对比及进化树分析
利用BlastP对SlETR6基因编码的氨基酸同源性进行检索,番茄SlETR6基因编码的氨基酸与花生AiETR2蛋白(XP_016175314.1)、向日葵HaETR2-like蛋白(XP_022002894.1)、胡桃JrETR2-like蛋白(XP_018810721.1)、苦瓜McETR2蛋白(XP_022146885.1)、木薯MeETR2-like蛋白(XP_021607133.1)、烟草NaETR2-like蛋白(XP_019227687.1)、樱桃PaETR2-like蛋白(XP_021807404.1)、矮牵牛PhETR蛋白(AAZ81985.1)、梅PmETR2(XP_008224393.1)、桃PpETR2蛋白(XP_007221962.1)、蓖麻RcETR2蛋白(XP_002529316.1)、芝麻SiETR2(XP_011091175.1)、马铃薯StETR-like蛋白(XP_006354517.1,)、枣ZjETR2蛋白(XP_015887972.1)等具有较高的相似性(图2)。系统进化树分析表明,其中番茄SlETR6与马铃薯StETR-like蛋白的同源性最高(图3)。

M. Marker Ⅲ DNA 分子标记;SlETR6. PCR 产物。M. Marker Ⅲ DNA Marker; SlETR6. PCR product.

表1 SlETR6启动子序列顺式元件分析Tab.1 The analysis of putative cis-elements in the promoter of SlETR6 genes


图2 番茄SlETR6与其他相似性高的氨基酸聚类分析Fig.2 Phylogenetic tree of amino acid sequences of SlETR6 of tomato and other similar amino acids

图3 番茄SlETR6 与其他植物氨基酸序列的进化树分析Fig.3 Phylogenetic tree of SlETR6 and other plant species
2.3 SlETR6基因的组织表达分析
qPCR检测发现,SlETR6在番茄根、叶、花、果实中均有表达,其在花中相对表达量较高,在叶中最低,在花、红果和叶片中的表达量与种子相比存在显著差异(图4-A);对SlETR6在果实不同发育时期的检测发现,转色果中表达量最高,红果和大果次之,在转色果、红果和大果中的表达量与小果相比存在显著性差异(图4-B)。
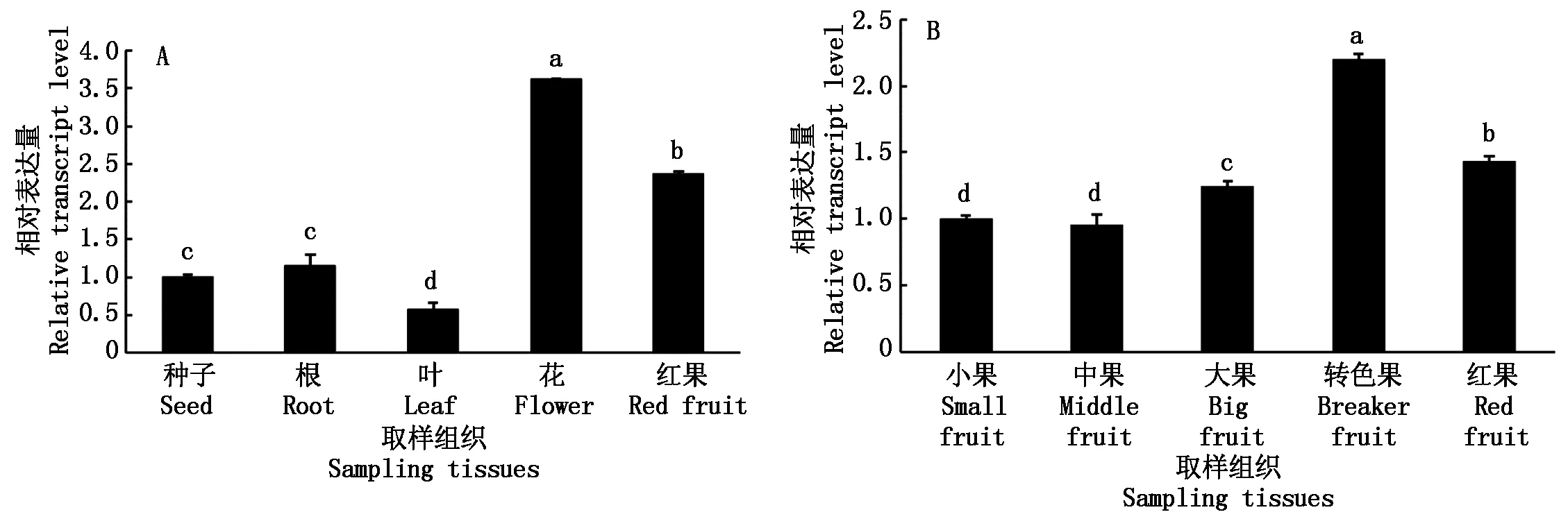
A.不同植物组织中SlETR6基因的表达情况;B.果实发育不同时期SlETR6基因的表达情况。不同小写字母表示在0.05水平差异显著。图5-7同。A. Relative expression of SlETR6 in different tissues; B. Relative expression of SlETR6 at different stage of fruit development. Different small letters show significantly different at 0.05.The same as Fig.5-7.
2.4 SlETR6基因在高盐胁迫下的表达分析
为了解SlETR6基因对高盐响应的表达特征,利用qPCR检测了其在胁迫后不同时间段的表达情况。结果显示:高盐胁迫处理6,12 h后SlETR6相对表达量具有显著性提高,且在处理后12 h表达量达到最高,相对表达量约为对照组的3.6倍,损伤处理24 h后SlETR6基因的相对表达量恢复正常水平(图5)。
2.5 SlETR6基因在温度胁迫下的表达分析
如图6所示,高温 (40 ℃) 储藏后SlETR6基因表达量表现出先升高(1~3 h)后下降(6~12 h)的趋势,并在高温处理3 h后达到峰值。高温胁迫24 h后其相对表达量恢复正常水平。低温4 ℃胁迫处理后,SlETR6基因相对表达量在低温处理后1,6,12,24 h表现出下调趋势。
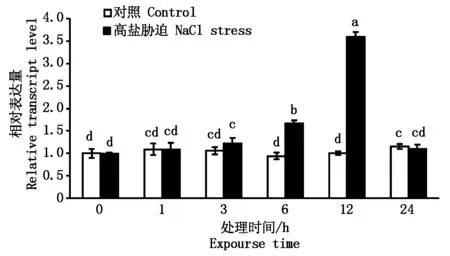
图5 高盐胁迫下SlETR6基因的表达情况Fig.5 Relative expression of SlETR6 under high salt stress

图6 温度胁迫下SlETR6基因的表达情况Fig.6 Relative expression of SlETR6 under temperature stress
2.6 SlETR6基因在干旱胁迫下的表达分析
在干旱胁迫条件下,SlETR6基因胁迫处理1 h后迅速应答,与对照组相比,其相对表达量提高1倍,3 h后其表达量呈下降趋势,但仍高于同期对照组。干旱胁迫处理24 h后,SlETR6基因的相对表达量恢复正常水平(图7)。与同期对照组相比,SlETR6基因在干旱胁迫的初期(1,3,6 h)均表现出显著性上调表达。

图7 干旱胁迫下SlETR6基因的表达情况Fig.7 Relative expression of SlETR6 under drought stress
3 结论与讨论
植物抗逆效应是植物对抗不良环境,比如抗旱、抗盐碱、抗涝、抗风、抗冻、抗病虫害等的能力。逆境的种类多种多样,主要可分为生物逆境和非生物逆境两大类。研究植物在非生物胁迫条件下的调节机制,对通过植物育种改变作物的特性、培育抗逆新品种有重要的意义。乙烯在响应植物逆境胁迫过程中有重要的作用[25-29],而对于乙烯受体在响应非生物胁迫过程中的研究较少,SlETR6是番茄乙烯受体家族成员之一,目前,尚无SlETR6参与非生物胁迫应答中的功能研究相关报道。
本研究在番茄中克隆了乙烯受体家族成员之一SlETR6,含有2 265 bp的开放读码框,编码754个氨基酸。乙烯受体是乙烯信号转导的负调控因子,参与了植物生长发育的多个过程。序列分析及同源性比对分析表明,SlETR6与马铃薯ETR-like蛋白的同源性最高。启动子序列分析表明,SlETR6具有多种逆境响应功能元件,如热胁迫、干旱胁迫、低氧胁迫、光响应、激素(乙烯、水杨酸、赤霉素)诱导响应、防御与胁迫响应元件等。
通过实时荧光定量PCR分析了SlETR6基因在种子、根、叶、花、花后5,10,15,25,35(绿熟期果实),45 d(红熟期果实)等不同组织中的相对表达量,SlETR6在番茄的不同组织中均有表达,且在花和转色期果实中表达量较高,推测SlETR6可能对于花的发育及果实的成熟过程有一定的调控作用。
另外,SlETR6对不同胁迫的响应模式不同。在4,40 ℃ 2个温度逆境胁迫环境下,SlETR6受到高温胁迫后强烈诱导,且在处理后3 h左右达到峰值,随后表达水平小幅下调,但仍然显著高于对照组,在处理后24 h恢复正常水平;高盐胁迫下,SlETR6在处理6 h后被诱导呈上调表达,并在处理后12 h达到峰值,为对照组的3.6倍;SlETR6在干旱胁迫早期受到强烈诱导,在处理后1 h达到峰值,是对照组的2倍。qPCR结果表明SlETR6同时受到高盐、高温、干旱等胁迫条件诱导,且生物信息学分析表明SlETR6基因启动子序列含有大量胁迫应答元件,包括干旱、热、防御反应相关的元件。综上,推测SlETR6基因在响应胁迫应答中发挥重要调控作用。本研究为后续研究SlETR6在响应逆境胁迫过程中的功能及作用机制奠定基础,为将来利用分子育种手段培育番茄抗逆新品种提供重要的基因资源。
猜你喜欢
做人与处世(2022年6期)2022-05-26
疯狂英语·新阅版(2022年5期)2022-04-29
中国特种设备安全(2022年1期)2022-04-26
当代工人·精品C(2019年1期)2019-04-29
中学生数理化·中考版(2016年7期)2016-12-07
汽车维修与保养(2015年8期)2015-04-17
高中生学习·高三版(2014年3期)2014-04-29
儿童故事画报·发现号趣味百科(2014年1期)2014-03-31
科技传播(2012年2期)2012-07-05
绿色科技(2010年8期)2010-08-15
