
马尾松人工林小气候调节效应
2019-03-06 01:50刘士玲杨保国雷丽群庞圣江韦菊玲
中南林业科技大学学报 2019年2期
刘士玲 ,杨保国 ,郑 路 ,雷丽群 ,庞圣江 ,张 培 ,李 华 ,韦菊玲
(1.中国林业科学研究院热带林业实验中心,广西 凭祥 532600;2.广西友谊关森林生态系统国家定位观测研究站,广西 凭祥 532600)
森林小气候具有协调生物与环境的关系的作用,是评价森林系统综合效益的重要特征之一[1]。无论是开展森林生物的生理生态特征、养分循环和水文功能等方面的研究,还是开展其他森林生态系统结构、功能等方面的研究,都不可缺少对森林小气候的观测[2]。传统的小气候研究注重对气象资料的简单描述,对小气候的时间和空间变异把握不足[3]。以往国内对森林小气候观测多为人工观测,局限于特定时间段的小气候特征分析,而缺少连续高频率的观测数据作为研究支撑,难以对森林的气候状况变化进行全面分析和把握。
马尾松Pinus massoniana是我国主要的用材与造林树种之一,分布区域广且适应性强。近年来,国内外关于马尾松的研究主要集中在材性[4]、种子园[5]、外生菌根真菌[6]、毛虫发生规律与防治[7]、地力维护[8]等方面,鲜有利用定点连续高频率的观测数据研究马尾松林小气候特征。本研究基于广西友谊关森林生态系统国家定位观测研究站的自动气象站对气象因子的高精度观测,以不同的尺度,对比马尾松林与空旷地气象因子差异,既可丰富关于马尾松生理生态方面的研究内容,也可揭示广西大青山地区不同下垫面水热的变化规律,同时也为本区域森林经营提供科学依据和基础数据。
1 研究区概况
研究区域位于广西友谊关森林生态系统国家定位观测研究站实验区范围内(106°41′~106°59′E,21°57′~ 22°16′ N),属南亚热带半湿润-湿润季风气候,有明显的干湿季。年均气温21.5~21.7℃,≥10℃积温6 000~7 600℃,年降水量1 200~1 500 mm,年蒸发量1 261~1 388 mm,相对湿度80%~84%。地貌类型主要是低山丘陵,海拔430~680 m。土壤为由花岗岩发育而成的赤红壤,土层厚度100~150 cm。
林内自动气象站安装在马尾松林,林龄为29 a,树高约19.1 m;灌木层以大沙叶Pavettaarenosa、古钩藤Cryptolepisbuchananii为主;草本层主要有铁芒萁Dicranopterisdichotoma、半 边 旗Pterissemipinnata、蔓生莠竹Microstegiumfasciculatum、弓果黍Cyrtococcum patens等。林外气象站放置在热带林业实验中心伏波实验场红星站(距马尾松林<1.5 km),根据国家气象局编制的《自动气象站建设技术要求》和《地面气象观测规范》建立。
2 研究方法
2.1 观测设备

表1 传感器的主要特征Table 1 Main characteristics of sensors
2.2 数据处理
采用气象划分法划分春季(公历3—5月)、夏季(公历6—8月)、秋季(公历9—11月)、冬季(公历12月—次年2月),且冬、春、夏、秋季的代表月份分别为1、4、7、10月。该区地处南亚热带,干湿季水热条件差异较大,因此选取干季1月份和湿季7月份分析林内外小气候日变化的差异。
统计14:00相对湿度≥80%和≤30%的出现日数,用于空气干、湿小气候特征的研究。以≥80%作为高湿指标,≤30%作为低湿指标[9]。
选取2016年全年数据进行分析。采用Microsoft Excel 2007对修订校正后的各要素的5 min步长观测数据加以整合,采用SigmaPlot 11.0完成图形分析。
3 结果与分析
3.1 小气候因子日变化规律
3.1.1 空气温度日变化
林内外气温日变化在1月(图1a)和7月(图1b)均呈现同步的倒U型曲线,7:00气温开始升高,15:00达到日气温最大值,随后逐步降低。林内日气温低于林外,1、7月份林内外气温日振幅分别为 2.25 ℃、3.00 ℃和 4.59 ℃、6.04 ℃。从干、湿季林内外气温差值变化趋势图(图1c)可以看出,相同时刻林内外差值表现为白天温差大于夜晚,湿季林内外气温差值大于相同时刻干季的差值,但波动幅度与峰值出现时间不尽相同。1月份林内外气温差值从6:00(0.55 ℃)的最低值开始上升,13:00(1.30 ℃)—16:00(1.35 ℃ )差值相似且为日差值最大的时段,随后16:00迅速减小至20:00(0.74 ℃)后林外气温差值曲线又趋于平稳,日平均温差为0.86℃。7月份差值曲线从7:00(0.69 ℃)的最低值开始上升,12:00达到最高温差2.17 ℃,随后16:00开始逐渐减小至21:00(0.74 ℃),日平均温差为1.34 ℃。
3.1.2 空气相对湿度日变化
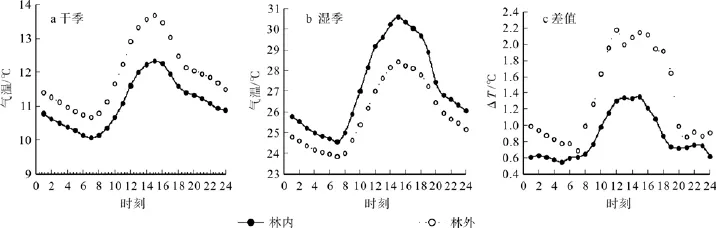
图1 空气温度日变化及差值Fig.1 Diurnal variations and differences of air temperature
空气相对湿度日变化在1月(图2a)和7月(图2b)均呈现同步的U型曲线,二者差值曲线同气温差值曲线相似,为倒U型曲线,峰值均出现在清晨时分,最低值在15:00—16:00,干湿季空旷地和马尾松林内相对湿度日振幅分别为8.54%、5.82%和29.00%、17.79%。干湿季空气相对湿度差值曲线变化趋势相似,湿度差白天大于夜晚,马尾松林内相对湿度均大于同时刻空旷地,湿季空气相对湿度差值曲线振幅大于干季,湿度差的较大值出现在10:00—19:00。干湿季日相对湿度差值最大值分别为 3.96% (13:00)、16.56(15:00),最小值分别为1.17%(8:00)、5.36%(7:00),平均差值分别为2.32%、10.42%。

图2 空气相对湿度日变化及差值Fig.2 Diurnal variations and differences of air relative humidity
3.1.3 土壤温度日变化
北方地区,早春及秋末气候较为冷凉,一般3月末至4有初在设施内育苗,终霜后定植于露地,6~9月份收获。因苦瓜种子种皮较厚,播前要进行浸种催芽。先用50~60℃的温水浸种,水冷后继续浸泡36~48小时,使其充分吸水膨胀。浸种后用纱布包好,放置于30℃左右的保温环境下催芽。将出芽后的种子直接播于营养钵内进行护根育苗。苗龄30~40天,2~3片真叶时移植。
干、湿季马尾松林内外土壤温度的时态变化趋势基本一致,但林内各层土壤温度稍低于林外空旷地。从季节上来看,湿季土壤温度的变化幅度较大。林内外土壤温度曲线最值出现的时间随土壤深度的增加而逐渐向后推移,即表现为迟滞效应(图3),且随土壤深度的增加土壤温度的变化幅度减小。5 cm土壤温度日振幅最大,干季林内外日振幅分别为0.50 ℃、0.58 ℃,湿季林内外日振幅分别为0.86 ℃、1.83 ℃,50 cm土壤温度日振幅近似为0。土壤温度的垂直变化表现为1月份随土壤深度的增加温度增加,7月份则相反。土壤温度差值曲线变化幅度表现为湿季>干季,干湿季土壤温度差值变化幅度均随土壤深度的增加而减小。
3.2 小气候因子月变化规律
3.2.1 空气温度月变化特征
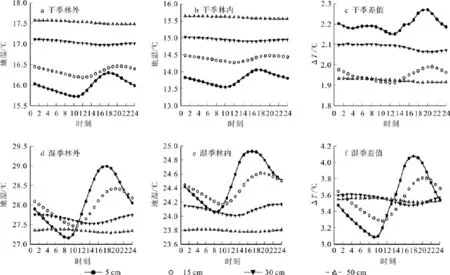
图3 土壤温度日变化规律及其差异Fig.3 Diurnal variations and differences of soil temperature
2016年马尾松林林内年平均气温为20.3 ℃,空旷地年平均气温为21.4 ℃,月气温差在0.8~1.3 ℃之间,二者差异不显著(P>0.05)(表1)。从季节上来看,马尾松林与空旷地的月均温差值表现出夏季>春季>秋季>冬季的规律,其中3至4月平均气温的变化幅度最大。从林内外空气温度月极值差异比较可以看出,林外空气温度月极端高温和极端低温均高于林内。林内外极端温度差异表现出一定的季节变化规律,即极端高温季节变化趋势为:夏季>春季>秋季>冬季, 林内外极端高温差异显著(P<0.05);极端低温季节变化趋势为:秋季>春季>夏季>冬季,其差异不显著(P>0.05)。马尾松林各月气温月均日较差均小于林外,且二者差异极显著(P<0.01),对比不同月份林分内外月均日较差的差值可以看出,其不同季节变化幅度大小为:夏季(2.03 ℃)>秋季(1.67 ℃)>春季(1.23 ℃)>冬季(1.13 ℃),夏季高温时马尾松林内温度相对林外较低,降温幅度较大,可有效地防止叶片灼伤;冬季低温时林内气温降低幅度变小,仅为夏季的55.6%,又起防寒作用,保障了马尾松林内稳定的气温变化,为林木生长提供了良好的条件。
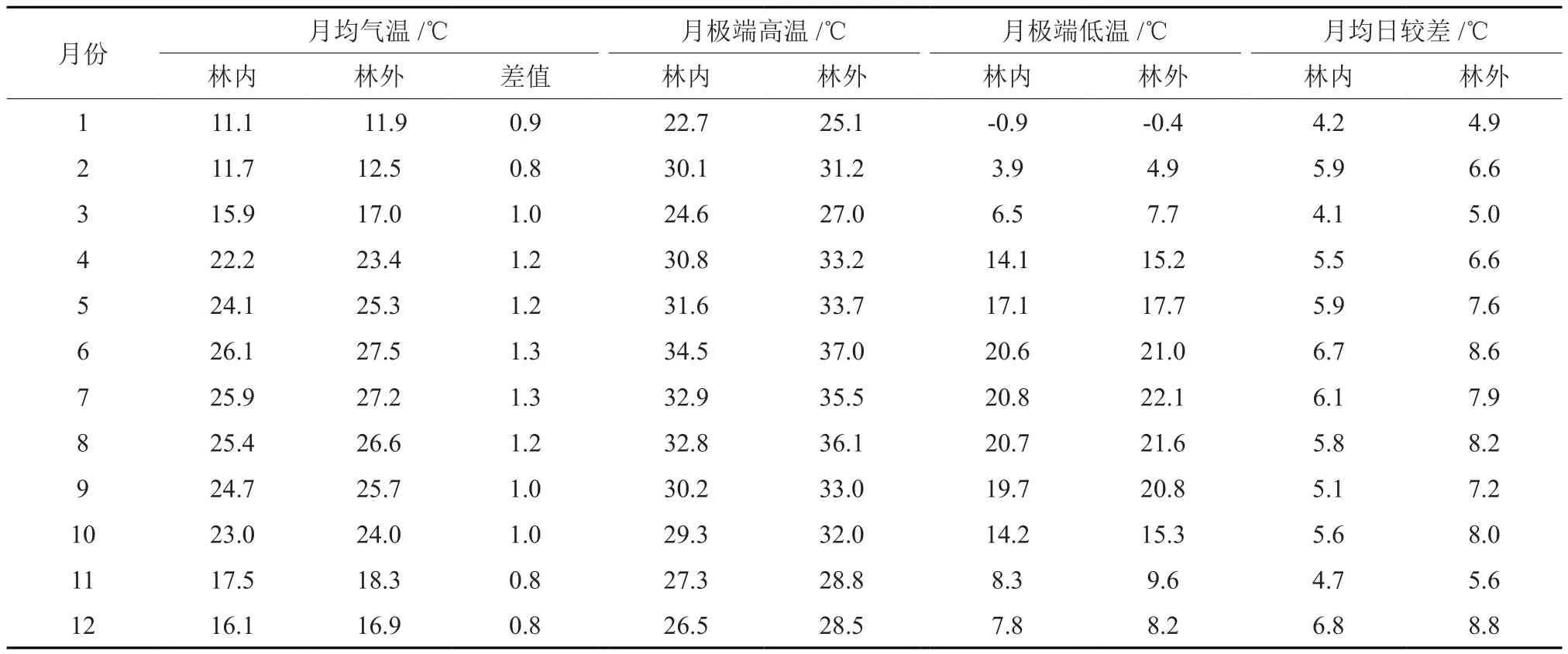
表1 马尾松林内外月气温比较Table 1 Comparison of monthly air temperature in forest area and open space area
3.2.2 空气相对湿度月变化特征
由表2可知,该区林内外相对湿度较大,均大于70%。马尾松林、空旷地年平均相对湿度分别为85.1%、80.4%,林内、外空气相对湿度差异性不显著(P>0.05)。通过统计14:00相对湿度≥80%和≤30%的出现日数发现,高湿日数林内多于空旷地,低湿日数林内少于空旷地。林内外不同季节空气相对湿度差值排序为:夏季>春季>秋季>冬季,这与空气温度差值季节变化规律相同,主要是因为生长季林木快速生长,蒸腾作用较强和林冠较大,对气流交换的阻挡效果较好,此外,春夏季雨水较多,使得夏春两季森林提高空气湿度的效果更为明显。
相对湿度林内较高,这是因为林木可以通过发达的根系从深层土壤吸取水分供林木蒸腾消耗,且林冠较大,对气流交换的阻挡效果较好,林地蒸发、植物蒸腾的水蒸气不易向外扩散,导致林内的空气湿度较大。
3.2.3 土壤温度月变化特征
由图4可知,马尾松林与空旷地土壤温度季节变化规律一致:夏季>秋季>春季>冬季。与日变化相似,随土层加深土壤温度变化幅度减小。林外空旷地各层土壤温度月振幅从14.1 ℃下降到12.1 ℃,马尾松林内则从12.0 ℃下降到10.3 ℃,由此可见,马尾松林具有较好的稳定土温的能力。林内外月均土壤温差为1.7~2.1 ℃,略高于月均气温差。林内外同层土壤温差年变化表现出先增大,6月份达到最大值后,又趋于减小,在12月份达到最小值,即干湿季差异更为显著(P<0.01)。
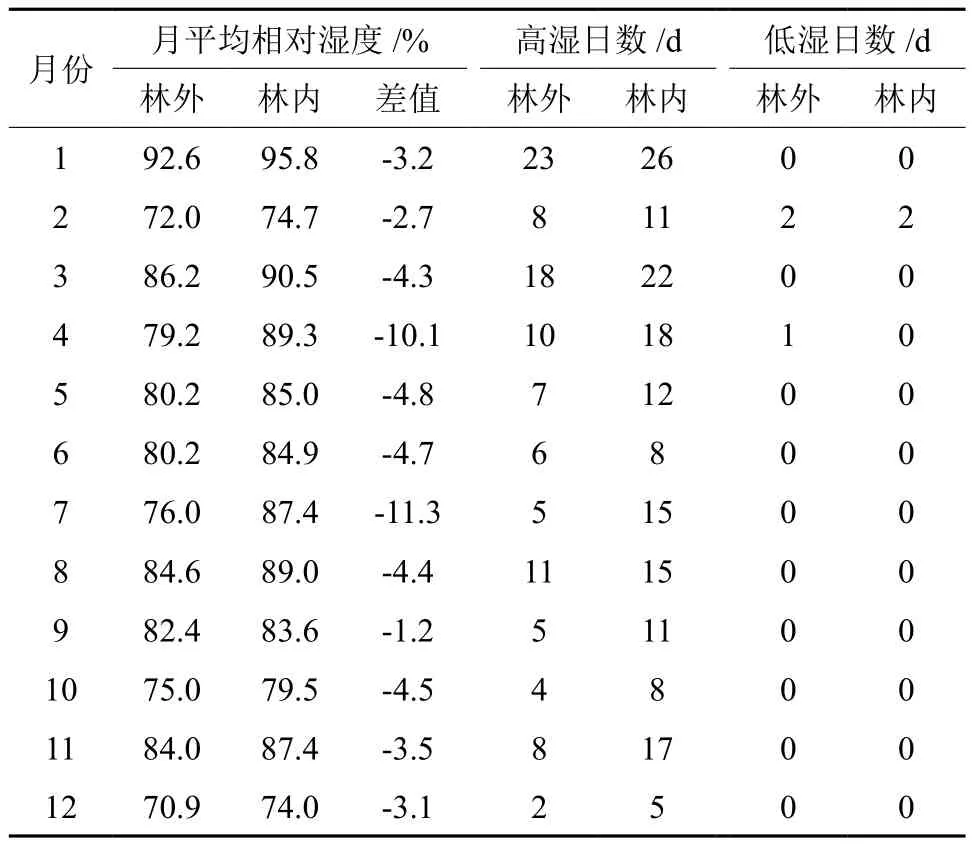
表2 林内外月相对湿度和高湿低湿日数的比较Table 2 Comparison of month relative humidity and number of days with high humidity and low humidity in forest and open space area

图4 各层土壤温度月变化Fig.4 Monthly variations of soil temperature at different soil depths
4 讨论与结论
4.1 结 论
1)林内外气温日变化在1月和7月均呈现同步的倒U型曲线,日气温林外高于林内。比较温度日较差可知,7月份林内外气温差值大于相同时刻1月份的差值,白天温差大于夜晚。从气温月变化来看,月均气温林外>林内,二者气温均呈现夏季>春季>秋季>冬季的规律。
2)1月和7月林内外空气湿度日变化为同步的U型曲线,二者差值变化为倒U型曲线,日振幅马尾松林大于林外。林内相对湿度均大于同时刻空旷地,林内年均空气相对湿度比空旷地高4.7%,湿度差均是白天大于夜晚。月均空气湿度林外<林内,从林内外空气湿度的季节变化规律来看,为夏季>春季>秋季>冬季。
3)干湿季林外与马尾松林内土壤温度的时态变化趋势基本一致,且林内外土壤温度曲线均表现出一定的迟滞效应,这种效应随土壤深度增加而逐渐减弱。土壤温度日变化和月变化均表现为林外各层高于林内,林内外同层土壤温差月变化表现出先增大后减小的趋势;从季节变化规律来看,马尾松林内外土壤温度均表现为夏季>秋季>春季>冬季。
4.2 讨 论
由于森林的存在使得林内与林外相比,热量交换在时间和空间上发生了显著的改变,从而使热量在林内重新分配,使林内温度发生了变化。本研究马尾松林年均温、年均湿度分别为20.3℃、85.1%,这与南亚热带鼎湖山处于中期演替阶段的的沟谷雨林小气候(年均温20.4℃、年均湿度87%)[10]极为相近,说明本研究马尾松林在改善森林气候功能方面已达到南亚热带鼎湖山演替中期阶段森林群落的水平,具有较好的调温保湿效应。沈运扩[11]研究表明,白蜡和怪柳人工林分别在春季和秋季具有保温作用,在夏季均表现为降温作用;蒋丹丹[12]通过对石灰岩山地两种模式人工林小气候比较研究,发现四季气温值均为林内<林外;这与本文马尾松林各月气温日较差均小于空旷地和四季气温均为林内<林外的研究结果一致,马尾松林夏季的降温作用更为显著,而冬季又能够一定程度上稳定林内温度,起到防寒作用。但这与周和锋等[13]杭州湾生态绿地小气候冬季林内温度高于林外的研究结果有所不同,这可能因为植被类型和研究区域地理位置的差异所造成的。
林内外空气湿度日变化在呈现同步的U型曲线,但不同生长季节的湿度最值和变化幅度有差异,这与闫文德和田大伦[14]研究的樟树人工林空气湿度的结果一致。闫俊华等[15]研究发现鼎湖山常绿阔叶林内外,空气相对湿度在春夏两季较大,秋冬两季较小,这与本文及徐云蕾等[16]林内外月均空气湿度差值排序夏季>春季>秋季>冬季的研究结果一致。马尾松林内外干季和湿季相对湿度差分别为10.42%、2.32%,相对于其他气候带而言,亚热带地区湿季的高温高降水将导致森林与空旷地的相对湿度差异变小,因此在水热资源相对缺乏的干季能表现出更好的增湿效应。从全年空气湿度来看,马尾松林高湿日数较多、低湿日数较少,表明马尾松林具有较好的保湿作用。
此外,由于小气候观测研究工作是一项长期的、系统的研究工作,今后应集中多年观测数据进行系统的研究,同时开展混交林与纯林小气候的对比研究、纯林及近自然化改造林分小气候的对比研究,更加全面观测与分析马尾松林小气候特征,从而为研究生态系统能量平衡、探讨生物与环境的关系打下良好的基础。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
西北林学院学报(2022年5期)2022-10-04
农业灾害研究(2022年1期)2022-05-07
土壤(2022年1期)2022-03-16
新农业(2021年12期)2021-11-29
棉花学报(2020年3期)2020-08-08
农家科技中旬版(2020年10期)2020-07-12
绿色科技(2019年5期)2019-11-29
江苏农业科学(2017年21期)2017-12-13
青年文摘·上半月(1988年2期)1988-11-01
