
干热河谷旱季3种典型植被恢复树种光合生理特征
2019-03-01 09:20韩学琴王雪梅邓红山廖承飞
西南农业学报 2019年1期
赵 广,金 杰,韩学琴,王雪梅,邓红山,廖承飞*
(1.中国科学院、水利部成都山地灾害与环境研究所,四川 成都 610041;2.中国科学院大学,北京 100049;3.云南省农业科学院热区生态农业研究所,云南 元谋 651399)
【研究意义】金沙江上游干热河谷地区植被破坏严重、水土流失加剧。近年来, 国家把长江中上游地区恢复植被作为生态防护林建设的重点不断加大投入,但蒸发大、降雨少的气候特点导致区域内生态恢复和重建工作开展极为困难[1]。人工栽种的植被恢复幼苗在旱季易受到土壤水分胁迫死亡,而如何在干热河谷筛选抗旱、速生以及适应能力强的植被恢复树种成为当前生态修复工作的重点和难点。光合作用是生态系统碳循环和水文循环的关键过程,植物光合生理对环境的适应性很大程度上反映了植物在该地区的生存适应能力和竞争能力[2],因此植物光合和水分生理特征往往作为植物逆境适应能力评价的重要指标[3]。探索植被恢复树种光合和水分生理特征可以更科学的做到适地适树,提高造林成功率,对干热河谷生态恢复有重要理论意义。但限于相关工作的局限性和阶段性,对干热河谷地区植物恢复树种光合和水分生理特征的比较和评价还很欠缺,以致难以科学指导生态恢复重建工作。罗望子(TamarindusindicaLinn)、银合欢(Leucaenaleucocephala)和辣木(MoringaoleiferaLam)是近些年元谋干热河谷的主要植被恢复树种,在当地生长良好。其中,罗望子又称酸角、罗晃子等,为苏木科(Caesalpiniaceae)罗望子属(Tamarindus)原产非洲,为热带亚热带常绿乔木型果树。主干明显,树体高大、抗旱、耐贫瘠,易于栽培管理,其果实酸角是糖、果汁、食醋等的优良原料。而银合欢是含羞草科(Mimosaceae)银合欢属的常绿乔木,原产美洲热带启动区,具有耐干旱、耐瘠薄、生长迅速等特点。银合欢繁殖扩散能力较强,可侵入其他郁闭度较大林分,有可能会对乡土树种生态系统产生生物入侵威胁[4],因此被列入外来入侵物种[5]。辣木又称鼓槌树,是辣木科辣木属热带落叶乔木,原产于印度北部,具有一定经济价值。辣木喜强光照,耐干旱贫瘠,适应通透性较好的砂壤土。【前人研究进展】目前,对干热河谷区罗望子和银合欢叶片光合生理特性的研究已有单独报道,但缺少树种间的对比;辣木的报道多集中于栽培技术、引种试种和产品开发等方面,缺少光合特性研究;整体上仍缺少对干热河谷植被恢复树种叶片水分利用效率的报道。【本研究切入点】本文以云南干热河谷地区罗望子、银合欢和辣木为对象,研究3种植被恢复树种光合参数对光强的响应特征,【拟解决的关键问题】探讨树种间的蒸腾和水分利用效率差异,为干热河谷进一步筛选植被恢复树种提供科学指导。
1 材料与方法
1.1 研究材料
研究区位于金沙江干热河谷地区元谋县境内(101°52′18″ E, 25°41′15″ N),地处滇中高原北部,隶属云南省楚雄彝族自治州。研究区属于南亚热带气候,具有干燥、炎热、少雨、光照充足、四季不分明而干湿季明显,全年无冬等特点。6-10月为该区雨季,11月至翌年5月为干季。全年降水量为613.9 mm,年蒸发量为3847.8 mm。试验区土壤以燥红土为主,干旱瘠薄,水肥条件极差。自然植被以草丛为主,杂以灌木,稀少乔木,称为半自然稀树草原或稀树灌草丛。
供试植株辣木、银合欢和罗望子均来自云南省农科院热区生态农业研究所辣木试验基地,均长势良好。
1.2 试验方法
试验于2017年8月下旬晴朗日子进行。选择生长健壮、长势一致3种植被恢复树种的成熟叶片,从当地时间9:00到12:00,采用便携式光合系统分析仪LI-6400(Li-Cor,Lincoln,NE,USA)分别测定辣木、银合欢和罗望子的Pn-PAR响应曲线。测定前叶片经过光诱导,用6400-02B LED光源将光合有效辐射(PAR)梯度设定为2000、1800、1500、1200、1000、800、500、250、100、50、0 μmol·m-2·s-1。测定同时记录气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和蒸汽压亏缺(Vpdl)等参数,每个树种测定3株作为重复。测定叶片均选取树冠中部南面一级侧枝健康、成熟叶片,测定时将叶片平铺于整个叶室,避免相互遮挡。光合测定结束后剪下叶室中的叶片,经扫描后用Image J图像分析软件进行叶面积校正,并根据实际叶面积计算出实际Pn、Gs、Tr、Ci和Vpdl等参数。
1.3 数据处理
本文采用非直角双曲线修正模型[6]拟合光合作用的光响应过程。
(1)
式中,Pn(I)为净光合速率,I为光合有效辐射(PAR),Ic为光补偿点,α、β、γ为独立于I的系数。
光饱和点(Isat):
(2)
暗呼吸速率(Rd):
Rd=-P(I=0)=-αIc
(3)
表观量子效率为I=Ic处的量子效率:
(4)
式中,Фc为表观量子效率(Apparent quantum yield,AQY)[7]。
叶片瞬时水分利用效率(Water use efficient,WUE)计算如下:
(5)
式中,Pn为叶片净光合速率,Tr为蒸腾速率。由于测得的光合和蒸腾均为瞬时速率,因此,(5)式计算出的WUE也被称作瞬时水分利用效率采用Excel 2007 处理实验,采用单因素方差分析法(One-way ANOVA)和最小显著差异法(LSD)比较树种间的光合参数差异,采用Origin 8.0完成绘图。
2 结果与分析
2.1 不同植被恢复树种主要光合参数对光强响应特征
由图 1可以看出,3种树种的Pn和Gs对PAR变化表现出不同的响应规律。弱光条件下(PAR<500 μmol·m-2·s-1),3种树种Pn随PAR增加迅速升高,但罗望子Pn升高速率较慢,辣木Pn增大速率最快。罗望子Pn在1000~2000 μmol·m-2·s-1范围内逐渐趋于平缓,银合欢Pn在1200~2000 μmol·m-2·s-1范围内趋于平缓,而辣木Pn则1500 μmol·m-2·s-1之后趋于平缓。整体上, 3树种的Pn表现为辣木>银合欢>罗望子。当PAR为0 μmol·m-2·s-1时,辣木和罗望子的Gs分别为0.0285和0.0184·H2O·μmol·m-2·s-1,而银合欢Gs则显著高于前两者(0.0720 μmol·m-2·s-1)。罗望子和银合欢Gs随PAR增强缓慢升高,而辣木Gs随PAR增强迅速升高,在1200~2000 μmol·m-2·s-1高于银合欢Gs。
当PAR变化0~500 μmol·m-2·s-1范围内,3个树种的Ci和Ci/Ca随PAR增强迅速降低;0~500 μmol·m-2·s-1范围内,Ci和Ci/Ca趋于稳定。整体上,3个树种的Ci和Ci/Ca随PAR变化不存在显著差异(P>0.05)。
2.2 不同植被恢复树种气体交换参数特征及相关关系
由表1显示,辣木Pmax(20.66 μmol CO2m-2·s-1)、Ic(125.24 μmol·m-2·s-1)和Rd(3.14 μmol CO2m-2·s-1)均显著高于银合欢和罗望子(P<0.05);银合欢的Pmax(10.05 μmol CO2m-2·s-1)、Isat(1741.87 μmol·m-2·s-1)、AQY(0.0369 mol·mol-1)和Rd(2.48 μmol CO2m-2·s-1)显著高于罗望子(P<0.05);辣木和银合欢的Isat、AQY差异不显著(P>0.05)。
由表 2显示,除罗望子叶片Pn和Vpdl的相关性未达到显著性(P>0.05),其他叶片Pn和环境因子的相关性均达到显著性水平(P<0.01);银合欢叶片Pn与PAR、Gs、Ci和Tr的相关系数高于辣木和罗望子;整体上,叶片Pn与PAR、Gs和Ci拥有较高相关性。
2.3 不同植被恢复树种蒸腾速率和水分利用效率特征
从图 2可以看出,3种树种的Tr和WUE对PAR变化表现出不同的响应规律。3树种叶片Tr
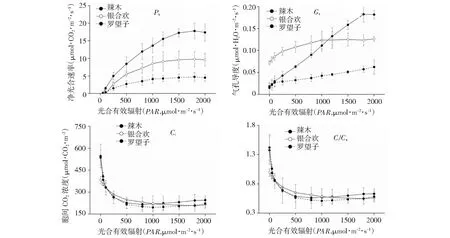
图1 3种植被恢复树种主要光合参数对光强的响应特征Fig.1 Responses of main photosynthetic parameters to PAR in three vegetation restoration trees

表1 3种植物叶片气体交换参数特征比较Table 1 Comparison of leaf gas exchange parameters of three vegetation restoration trees
注:表中字母为最小显著差数法(LSD)进行的多重比较结果,小写字母代表0.05水平下的检验结果。
Note: The letters in the table are the result of multiple comparisons by the least significant difference method(LSD), and the lowercase letters represent the test results at 0.05 level.

表2 3种植物叶片净光合速率与其他因子的相关关系Table 2 Correlation coefficients of three vegetation restoration trees between Pn and other factors
注:**在0.01水平(双侧)上显著相关。
Note:**represents signifance at 0.01 level (two-tailed).
都表现为随PAR增强而逐渐升高,整体大小表现为银合欢>辣木>罗望子;所有树种的WUE在0~500 μmol·m-2·s-1范围内随PAR增强急剧升高,500~2000 μmol·m-2·s-1范围内趋于稳定,3树种的WUE整体表现为辣木>罗望子>银合欢。
3 讨 论
3.1 3种植被恢复树种主要光合参数对光强响应特征
光合作用是植物体内重要的代谢过程,其强弱很大程度上决定了植物生长发育和抗逆特性,因而往往被作为判断植物生长状况的重要指标[8]。本文中,相同有效光合辐射下,辣木具有较高的Pn,其次是银合欢和罗望子,表明辣木具有较强的光合同化速率和生长适应能力。作为速生乔木,辣木一年生长高度能达到6~7 m[9]且当年结果,这种快速的同化产物积累必然要以较强的光合同化能力为支撑;而罗望子生长速率相对缓慢,8~12年才能结果,可能与其较弱的光合同化能力有关。
气孔是植物叶片与外界进行气体交换的主要通道,其调节能力的有效程度是植物适应不同环境的重要方式[10]。气孔开闭是迅速、灵敏可逆的生理过程[11],气孔导度对光强的响应反应了植物叶片对不同光质量的利用状况。银合欢和罗望子叶片气孔导度随有效光合辐射增强而平稳上升,而辣木叶片气孔导度则迅速升高。辣木有较强的光合同化能力,表明辣木叶片在强光环境下能迅速打开气孔、提高光合速率。

图2 3植被恢复树种叶片蒸腾速率和水分利用效率对光强的响应特征Fig.2 Responses of Tr and WUE to PAR in three vegetation restoration trees
3.2 3种植被恢复树种气体交换参数特征及相关关系
光补偿点和光饱和点分别反映了植物叶片对弱光和强光的利用能力。研究中,辣木和银合欢具有较高的光饱和点,表明二者对强光具有较强的利用能力;银合欢和罗望子光补偿点较低,说明其能有效利用弱光。表观量子效率(AQY)表示植物吸收和转换光能的色素蛋白复合体的数量,尤其是其利用弱光的能力,与其叶片光能转化效率正相关[12]。正常生长的植物表观量子效率通常在0.03~0.05之间,数值越高表示叶片对光能的转化效率越高[13]。本研究中,3树种表观量子效率(AQY)总体上处在正常范围内,但辣木和银合欢的表观量子效率显著高于罗望子,这表明辣木和银合欢对干热河谷特殊环境具有较强的适应能力。
净光合速率(Pn)和其他因子的相关关系显示,光合有效辐射(PAR)、气孔导度(Gs)和胞间二氧化碳浓度(Ci)是3树种叶片光合作用的共同限制因子。胞间二氧化碳浓度(Ci)与净光合速率(Pn)呈负相关系,光合有效辐射(PAR)和气孔导度(Gs)与与净光合速率(Pn)呈正相关系。光合有效辐射(PAR)增加会促进叶片升高气孔导度(Gs),增大净光合速率(Pn)。光合速率升高会增加CO2消耗量,导致胞间二氧化碳浓度(Ci)浓度降低,反之亦然[14]。
3.3 3种植被恢复树种蒸腾速率和水分利用效率特征
蒸腾耗水是植物光合、蒸腾作用的综合反映,是评价植物生长适应性的重要指标。作为叶片光合与生理过程的耦合因子,水分利用效率(WUE)是植物能量转换的一个重要指标,已成为干旱半干旱地区农学家和生态学家研究的热点问题[15]。本研究中,罗望子的叶片蒸腾速率显著低于银合欢和辣木,且对光合有效辐射变化表现出稳定性,表现出蒸腾特性对干热河谷地区强光照环境的适应性。罗望子叶片相较银合欢和辣木具有较大叶厚度和革质特性,而叶厚度增加能有效防止水分过度蒸腾[16],这可能是罗望子蒸腾速率较低的原因。
胡红玲等[17]研究发现,巨桉与其他5个树种的净光合速(Pn)和蒸腾速率(Tr)均随光强的增大(200~1500 μmol·m-2·s-1范围内)而升高,而WUE在光强800 μmol·m-2·s-1时均为最高,在光强1500 μmol·m-2·s-1时有所降低。这可能是因为在低光强范围内,光强增大导致光合速率的增幅大于蒸腾速率的增幅,而在较高光强下则相反,即过高或过低的光强都会降低植物水分利用效率的结论。而本文中,3种植物的叶片水分利用效率在有效光合辐射500~2000 μmol·m-2·s-1范围内变化不大,表现出植物特有的对干热河谷强光环境的适应性。辣木具有较强的光合产物同化能力和较低的蒸腾速率,因此叶片水分利用效率最高;罗望子叶片净光合速率虽然最低,但蒸腾速率处于低水平,反而水分利用效率高于银合欢;银合欢光合能力较高,但其生长策略是以高蒸腾速率和低水分利用效率为代价获取充足的同化产物。
4 结 论
3种树种的Pn和Gs对PAR变化表现出不同的响应规律;弱光条件下(PAR<500 μmol·m-2·s-1),3树种Pn随PAR增加迅速升高;辣木和银合欢具有较高的光饱和点,而银合欢和罗望子光补偿点较低;3树种表观量子效率(AQY)总体上处在正常范围内,但辣木和银合欢AQY显著高于罗望子(P<0.05);罗望子的叶片蒸腾速率(Tr)显著低于银合欢和辣木(P<0.05),3树种的WUE表现为辣木>罗望子>银合欢。整体上,辣木光合能力和水分利用效率较高,且生长迅速,而罗望子虽然光合速率较低、生长缓慢但经济利用价值最高,二者可考虑作为植被恢复树种推广研究;而银合欢具有高蒸腾、低水分利用效率的光合生理特征及其潜在的入侵植物危害属性,因此今后的干热河谷植被恢复物种筛选应谨慎选择银合欢。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
河南水利与南水北调(2022年1期)2022-03-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
福建农林大学学报(自然科学版)(2021年1期)2021-01-22
中国农业科技导报(2020年12期)2020-03-15
亚热带农业研究(2019年2期)2019-09-02
世界热带农业信息(2019年9期)2019-01-05
今日农业(2019年14期)2019-01-04
