
水稻纹枯病菌侵染稻株对褐飞虱发生的影响
2019-03-01 03:42张珏锋夏乐舜李芳钟海英陈建明
中国稻米 2019年1期
张珏锋 夏乐舜 李芳 钟海英 陈建明
(1浙江省农业科学院植物保护与微生物研究所/浙江省植物有害生物防控重点实验室省部共建国家重点实验室培育基地,杭州310021;2浙江丽水出入境检验检疫局,浙江丽水323000;第一作者:zhangjuefeng@sina.com)
褐飞虱(Nilaparvata lugensStl)是亚洲水稻产区的一种主要迁飞性害虫,以吸食水稻韧皮部汁液为食,易造成水稻倒伏、虱烧等现象[1]。水稻纹枯病(Rhizoctonia solaniKuhn)由立枯丝核菌引起,使水稻不能抽穗或抽穗后秕谷较多,粒重下降,严重影响水稻的产量[2]。褐飞虱与水稻纹枯病的发生时间、发生环境和发生部位基本相似,均在夏末秋初高湿的气候环境中为害或感染水稻植株叶鞘。由于褐飞虱分泌的蜜露促使纹枯病菌丝的生长和菌核的形成,且蜜露中所含氨基酸对增加病原菌的菌核数量和增加致病力有着极有利的作用[3],早期的田间调查也发现,稻飞虱发生严重的田块,纹枯病发生也相对较重[4-5],并且褐飞虱数量与纹枯病的发生呈极显著的正相关[6]。
由于植物在生长过程中受到昆虫为害或病原菌感染后,会通过产生萜类、生物碱、类黄酮等次生代谢物来抵抗入侵生物[7-8];自然界中通常会遭遇多种生物为害,病虫会同时为害一种植物,病害或虫害诱导的植物化学防御会产生一定程度的交互作用[9]。通常植物对病害和虫害的防御方式之间存在着相互抑制的拮抗作用,即发生虫害的植物更容易感病,发生病害的植物则更容易感虫[10]。褐飞虱与水稻纹枯病在发生条件、发生部位与发生时间上都存在重合。也有田间数据表明,褐飞虱应该更趋向于取食感染纹枯病的稻株,但目前缺乏纹枯病菌侵染水稻植株后对褐飞虱发生为害的影响研究,这将严重阻碍进一步阐明水稻褐飞虱的发生规律和灾变机制,并影响到科学合理制订水稻褐飞虱和纹枯病的复合防治指标,不利于稻田病虫害的合理防控。本文利用刺吸电位图(EPG)以及褐飞虱蜜露的测定等方法,结合研究不同抗虫水稻植株感染纹枯病菌后褐飞虱的取食行为变化,进一步明确水稻褐飞虱与水稻纹枯病的协同发生作用,为合理制订水稻褐飞虱和纹枯病的复合防治指标提供理论依据。
1 材料与方法
1.1 试验材料
供试水稻品种:感虫品种Taichung Nativel(TN1),抗虫品种Rathu Hennati(RH)。
供试褐飞虱:褐飞虱虫源采集于杭州田间种群,于浙江省农业科学院温室越冬用TN1稻苗繁殖2代后,孵化48 h内成虫进行试验。
供试菌株:水稻纹枯病菌Rhizoctonia solani由浙江省农业科学院植物保护与微生物研究所真菌实验室提供。
仪器:DC-EPG型四通道面板控制昆虫电子取食监测仪,荷兰瓦赫宁根农业大学研制。
1.2 试验方法
水稻纹枯病菌的接种:TN1和RH在30 d苗龄时,稻株基部接入纹枯病菌核,透明塑料容器罩住水稻苗,置于28℃±2℃,光照16 h∶8 h恒温培养箱中培养72 h,以未接纹枯病菌的水稻苗作为对照。
取食行为测定:参考何月平等[11]的方法,采用昆虫电子取食监测仪刺探电位图(electrical penetration graph,EPG)进行褐飞虱取食行为监测。将褐飞虱成虫分别置于健康的TN1稻株、RH稻株和感染纹枯病的TN1稻株、RH稻株上,每种处理重复20次,每个重复连续记录8 h。所有EPG试验在温度(26±2)℃、湿度(85±5)%的人工气候室中进行。采用随机附带的ACQ和ANA软件对褐飞虱取食行为进行记录和数据的统计分析。
蜜露量及蜜露氨基酸含量测定:Parafilm膜制作口袋,固定于4株不同处理的水稻苗基部,每株苗上放置5个Parafilm口袋,每个口袋内均放1头短翅雌成虫的褐飞虱,24 h后按黄建利等[12]的方法取出蜜露称量,并将收集的褐飞虱蜜露按傅强等[13]的方法于40℃条件下吹干,加900 μL 2%的磺基水杨酸水溶液溶解游离氨基酸后送测。

表1 不同处理下褐飞虱取食行为参数比较
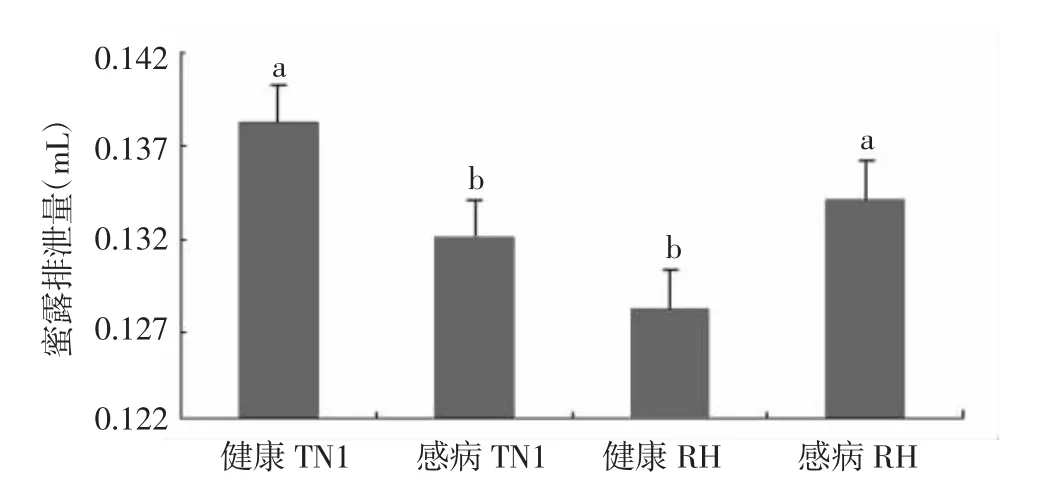
图1 褐飞虱取食不同处理水稻24 h后排泄的蜜露量比较
2 结果与分析
2.1 褐飞虱在感虫品种TN1和抗虫品种RH上的取食行为比较
褐飞虱在取食不同处理的抗感水稻品种过程中的EPG参数(表1)显示,试验过程中记录到np(非刺探)、N1(开始刺探)、N2(口针朝维管束移动的途中)、N3(口针在临近韧皮部外面移动)、N4-a(口针在韧皮部内部的一种活动)、N4-b(口针在韧皮部内稳定摄取汁液)、N5(口针在木质部摄取水分)共7种典型的褐飞虱取食波形。褐飞虱在健康TN1植株上的N1波形持续时间为28.16 sec显著低于其余3个处理,但在感病TN1植株上N1波形出现的次数显著少于RH植株的2个处理;N2波形在RH 2个处理上的出现次数及持续时间均显著高于感虫品种的处理;BPH在感病RH植株上的N3波形的持续时间显著长于其余3个处理;同时褐飞虱在RH植株的2个处理上,N2+N3波形的持续时间显著高于TN1的2个处理,但是N4波形在TN1植株上持续时间的百分率却显著高于RH植株处理。由此可知,感染纹枯病与否对褐飞虱在抗感水稻品种上的取食行为并无显著影响,褐飞虱在感虫品种TN1上韧皮部取食的刺探次数要显著少于其余处理。
2.2 褐飞虱在不同处理水稻上取食之后的蜜露排泄量及蜜露氨基酸含量比较
从图1可见,取食健康TN1植株的褐飞虱分泌的蜜露量为0.138 mL,显著高于取食感病TN1植株和健康RH植株的蜜露分泌量,但与取食感病RH植株的褐飞虱蜜露分泌量无显著性差异。从蜜露分泌的结果可推测,感染纹枯病的抗虫水稻品种比未感病品种更易吸引褐飞虱取食。取食不同处理水稻品种褐飞虱分泌的蜜露中多数必需氨基酸的含量并无显著差异,但取食感病TN1稻株的褐飞虱和取食感病RH植株的褐飞虱分泌蜜露中的甘氨酸(Gly)含量存在显著差异;同时,取食健康RH植株的褐飞虱分泌蜜露中的苯丙氨酸(Ile)含量和赖氨酸(Lys)含量均显著高于其余3个处理。
3 讨论
病原菌侵染引发植株防卫基因表达的信号传导途径是水杨酸途径(SA)[14],水杨酸途径与虫害(机械损伤)诱发的植物防御信号传导途径茉莉酸途径(JA)[15-16]会同时存在,且两种信号途径间存在相互抑制的拮抗作用[17-18]。本试验结果表明,水稻植株受到纹枯病菌侵染之后,在取食行为、蜜露排泄量及蜜露中氨基酸含量方面均未发现褐飞虱对感染纹枯病的抗、感水稻品种有显著地取食趋性。本次试验结果并未说明抗、感褐飞虱的水稻品种感染纹枯病之后更易感虫的性状。

表2 不同处理下褐飞虱体内必需氨基酸含量的比较 (nmol/mg)
Felton等[10]研究发现,烟草植株中苯丙氨酸解氨酶(PAL)基因的过量表达使植株对病原菌抗性提高,但对植食性昆虫的抗性下降,而不表达型植株的表现刚好相反,同时过量表达型烟草植株体内SA水平提高的同时,JA水平下降,说明PAL在水杨酸合成途径中起关键作用。纹枯病抗性不同的水稻品种接种水稻纹枯病菌后,稻株内PAL酶活性均有升高,但不同品种的酶活升高及持续的时间不同,有24 h内即开始升高,48 h后达到高峰后开始下降,72 h后即回落到初始水平;也有PAL酶活性水平在接种后有一定降低,48 h后急剧升高,并在第3 d和第7 d有2个高峰[19]。这说明不同水稻品种接种纹枯病菌之后体内PAL的生理变化机制是不同的。本试验中采用接种纹枯病菌72 h之后的抗、感褐飞虱水稻植株用于试验,植株均未表现出感虫症状,可能与所用水稻品种体内的PAL活性水平在该时间内并未显著升高有关。
此外,纹枯病菌菌丝体侵入稻株表皮后在受侵细胞内呈网状扩展,并穿透毗邻细胞壁,感病组织出现细胞变形、变空、腐烂等症状[20]。纹枯病菌能使接近菌丝的质膜发生质壁分离、质膜断裂、叶绿体变形、类囊体被破坏、线粒体解体等一系列组织病变[21]。而稻株组织内的病变可能对于在水稻韧皮部组织内取食稻株汁液的褐飞虱有较大的影响,即使稻株对褐飞虱的防御性降低,褐飞虱在对RH的感病植株和健康植株的选择中明显倾向于选择感染纹枯病菌的水稻植株,但稻株内部组织的破坏却阻止或减少褐飞虱刺吸性取食行为的产生。因此,可针对不同抗性水平水稻品种感染纹枯病菌之后体内PAL活性及基因表达量,以及稻株感染纹枯病菌之后对于水稻田内咀嚼式口器害虫(如二化螟、稻种卷叶螟等)的影响开展进一步研究。
猜你喜欢
上海农业科技(2022年4期)2022-08-15
华南农业大学学报(2022年5期)2022-08-02
热带作物学报(2022年5期)2022-06-06
热带农业科学(2022年5期)2022-06-02
特种经济动植物(2021年12期)2021-12-13
昆虫学报(2021年8期)2021-09-06
清风(2020年12期)2020-09-10
农业机械学报(2020年2期)2020-03-09
意林原创版(2019年2期)2019-03-04
椰城(2018年9期)2018-11-15
