
水分亏缺冬小麦近等基因系冠气温差与群体总耗水量的关系
2019-02-21 02:46梅旭荣黄桂荣严昌荣刘晓英张欣莹王雅静顾峰雪钟秀丽
生态学报 2019年1期
梅旭荣,黄桂荣,严昌荣,刘晓英,张欣莹,王雅静,顾峰雪,郭 瑞,钟秀丽
中国农业科学院农业环境与可持续发展研究所,国家作物高效用水与抗灾减损工程实验室/农业部旱作节水农业重点开放实验室, 北京 100081
在水资源危机,干旱化趋势不断加剧,以及由此引发的环境问题日趋严峻的背景下,选用高产甚至中等产量的低耗水品种,无疑是有效减少灌溉水量,缓解水资源短缺对农业生产影响的重要途径。研究品种之间耗水特性的差异及其关键影响因素,是找到品种耗水特性评价与节水品种鉴选方法的突破口。
已有大量研究从叶片与个体水平揭示不同品种间蒸腾耗水特性的差异与机理[1-2]。然而,群体水平的总耗水量受整个生育期内包括品种蒸腾生理特性等诸多因素的影响。因此,研究品种群体水平全生育期耗水量的差异及关键影响因素,对于品种耗水特性评价以及节水品种鉴选方法更有意义。群体水平的蒸腾与蒸发量不仅决定于空气动力学阻力和气孔阻力,还在很大程度上受控于冠层阻力[3-4]。群体内部因植株密度、高度、茎叶形状、数量和分布不同形成特定的冠层结构[5-6]。冠层结构作为植物的受光结构,不仅影响光的截获,还影响冠层与大气之间水热的垂直交换过程,从而影响冠层内部温度与湿度,并限制群体的蒸发与蒸腾耗水[7-9]。
冠气温差(Canopy-air temperature difference, CTD)即作物群体的茎、叶表层温度的平均值与周围大气之间的温度差值,反应植株内部生理代谢功能和冠层结构特性,受品种特性、冠层阻力、土壤水分条件、及饱和水蒸气压、大气温湿度、净辐射等环境因素的影响[10-12]。张嵩午等[13-14]最早关注冠层温度,并根据冠层温度将小麦品种分为冷型、暖型和中间型3种类型。大量研究发现品种间冠层温度的差异与经济产量[15-18]和抗旱性[19-21]密切相关。目前,冠层温度已被认定为品种抗旱性与产量潜力评价的重要依据。在大气因子与土壤水分条件相同的情况下,品种间CTD与群体总耗水量的差异均受控于品种特性与冠层特性,那么二者之间的关系如何,至今未见报道。
本研究选用一组近等基因系材料,其亲本品种晋麦47和京411,是从大量冬小麦品种中筛选出的CTD具极显著差异的材料。而且,在水分亏缺及灌溉条件下,两个亲本之间产量均没有显著差异,而耗水量达到极显著差异。它们产生的近等基因系因而是研究群体耗水量差异与机理的理想材料。本研究利用这组近等基因系,在水分亏缺条件下,设定种植密度与其他栽培措施一致,探讨群体冠层特性相关指标CTD及品种生理特性指标叶片蒸腾速率和气孔导度与群体耗水量的关系,旨在为探讨冬小麦材料耗水特性评价指标与低耗水品种的筛选方法提供依据。
1 材料与方法
1.1 试验材料及设计
供试材料为中国农业科学院作物科学研究所景蕊莲提供的冬小麦品种晋麦47、京411及其近等基因系15个。供体亲本为京411,轮回亲本为晋麦47,回交至BC3F4。
试验于河北省农林科学院衡水旱作节水农业试验站(37°54′N,115°42′E)进行。衡水试验站地处黄淮平原黑龙港地区,年平均气温13.0℃,无霜期206 d。2012年10月至2013年6月,生育期内平均气温为8.06℃。全年自然降水总量为491 mm,主要集中在6—8月,占全年总降水量的74%,生育期内降水量仅为177.9 mm。年日照时数2043 h。2016年10月到2017年6月,生育期内平均气温为11.9℃。全年自然降水总量约为537 mm,主要集中在6—8月,占全年总降水量的75%以上,生育期内降水量仅为125.2 mm。年日照时数2294 h。
试验条件设定为水分亏缺条件。用带有半自动防雨棚的防雨池开展,防雨池面积为3 m×2.2 m,池深3 m,土层深2 m,土壤类型为潮土质,下层1 m是粗砂渗滤层。在播种前灌底墒水,灌溉量以60 mm为标准,根据土钻法测定的土壤含水率调整实际灌量,使每个小区土壤含水量一致。各小区随机设计,3次重复。于2012年10月20日播种,2013年6月5日收获。2016年10月15日播种,2017年6月7日收获。行距20 cm,播种密度为3.3×106株/hm2。两个年度分别在干旱严重的拔节初期即2012年4月5日和2016年4月6日补充灌溉一次,根据每个小区监测的土壤含水率计算灌溉量,使得每个小区的土壤含水量能达到田间持水量的65%。除水分管理外,其他均为常规管理措施。
1.2 测定指标与方法
1.2.1 冠层温度和空气温度测定
采用手持式德产testo 845红外测温仪,在冬小麦的拔节-孕穗期、抽穗-开花期和灌浆期,分别选择2013年4月17日、5月3日和5月22日,2017年4月15日、5月2日和5月23日,3个晴朗无风的天气,于13:00—15:00进行测量。为消除太阳方位角和种植方向对观测值的影响,测定时仪器探棒沿小区种植走向, 在小麦冠层上方垂直距离15—20 cm,仪器与作物冠层呈30°夹角,测定生长一致、有代表性的部位,避免红外线照射裸露地面,直接读出冠层温度值和空气温度值。每小区测3次,取平均值作为该次测定的冠层温度值和空气温度值,计算二者差值,即为冠气温差CTD。
1.2.2 叶片蒸腾速率和气孔导度的测定
选取受光方向和生长状况一致的叶片3片,采用美国LI-COR公司生产的LI-6400便携式光合作用测定仪,分别于2013年4月15日、5月1日和5月20日和2017年4月13日、4月30日和5月18日9:00—11:00测定叶片蒸腾速率与气孔导度。
1.2.3 土壤水分监测与耗水量测定
在播种前和收获后用土钻取土法测定土壤含水率。冬小麦返青后每隔10 d采用TRIME时域反射仪监测土壤体积含水率,测深160 cm,20 cm一个层次。因为有防雨棚不受自然降水的影响,生育期耗水量(ET)=灌水量+播种时土壤含水量-收获时土壤含水量-池底渗水量。本研究实验条件下,池底渗水量均为0。
1.2.4 产量测定和水分利用效率计算
成熟后,对3 m×2.2 m小区采取人工镰割方式收获,装于网袋,自然晒干后人工脱粒称重,折算每公顷产量(Y)。水分利用效率WUE=供试品种籽粒产量(Y)/供试品种耗水量(ET)。
1.2.5 数据处理
应用SAS(Statistical Analysis System)9.0软件进行CTD方差分析和材料之间各种性状的差异显著性分析。
2 结果与分析
2.1 土壤水分的变化动态
实验于自动防雨棚和防雨水分栽培池中开展,因而没有自然降水,只在播种前灌底墒水,在干旱严重的拔节初期补充灌溉一次。由图1可见,2012—2013年度和2016—2017年度栽培池不同深度土壤水分动态变化趋势非常相近,从播种前到收获之日土壤体积含水率一直处于下降状态,4月中旬出现一个小高峰,是因为4月初补灌的结果。播种前各层土壤体积含水率2012—2013年度为31.4%—35.2%,2016—2017年度为28.3%—33.1%。收获时各层土壤体积含水率2012—2013年度为13.3%—23.0%,2016—2017年度为12.0%—20.7%。不同土层土壤水分下降速度不同,0—20 cm土层下降最快,因而下降幅度最大,生育期内土壤含水率明显低于下面土层。
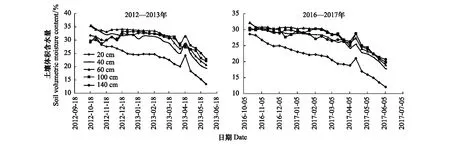
图1 不同深度土层土壤体积含水率的变化Fig.1 Variation in soil volumetric moisture of different soil depth
2.2 不同冬小麦近等基因系CTD的差异
2012—2013年度,拔节-孕穗期、抽穗-开花期和灌浆期3个不同生育期,15个近等基因系及其亲本的CTD平均值分别为7.5,6.19、5.68℃。2016—2017年度,3个生育期的CTD平均值分别为6.25、5.9、4.77℃。两个年度CTD的变化趋势一致,均是随着发育进程推进,虽然气温逐渐上升,但是冠层与周围大气之间的差值CTD是降低的趋势(表1)。3个生育期部分近等基因系及其亲本之间CTD存在显著差异,供体亲本京411最低,而晋麦47最高。从CTD最高的拔节-孕穗期来看,2012—2013年度,CTD最低的一组材料为京411、907990和908054,CTD最高的一组材料为晋麦47、 908188、 908106、 908120和907984。 2016—2017年度的结果与2012—2013年度的结果趋势非常相近,只是略有差异。拔节-孕穗期,京411的CTD与907990和908054虽有显著差异,但三者均排在后三位,所以总体趋势没有差别。
CTD的方差分析表明,基因型和重复均对CTD有显著影响,但是二者之间只有在抽穗-开花期存在互作(P=0.0002),而在拔节-孕穗期和灌浆期均没有互作(表2)。随着发育进程推进,基因型对CTD的影响减弱,拔节-孕穗期,品种/系之间CTD差异达到极显著水平,而抽穗-开花期和灌浆期,基因型之间差异减小,但均达到极显著水平。
2.3 不同冬小麦近等基因系总耗水量的差异
水分亏缺条件下,15个近等基因系及其亲本京411和晋麦47之间耗水量存在显著差异(表3)。2012—2013年度,京411与908240、907986、908092和908054的耗水量为最高一组,显著高于晋麦47,另外11个系与晋麦47没有显著差异。晋麦47与908188、90274、908120、908106、908216、907988、908176和908174的耗水量为最低一组,显著低于京411,另外7个系与京411没有显著差异。2016—2017年度,耗水量最高的一组为京411、908092和908054,耗水量最低的一组为晋麦47、908188、90274和908174。两个年度的实验结果趋势相近。
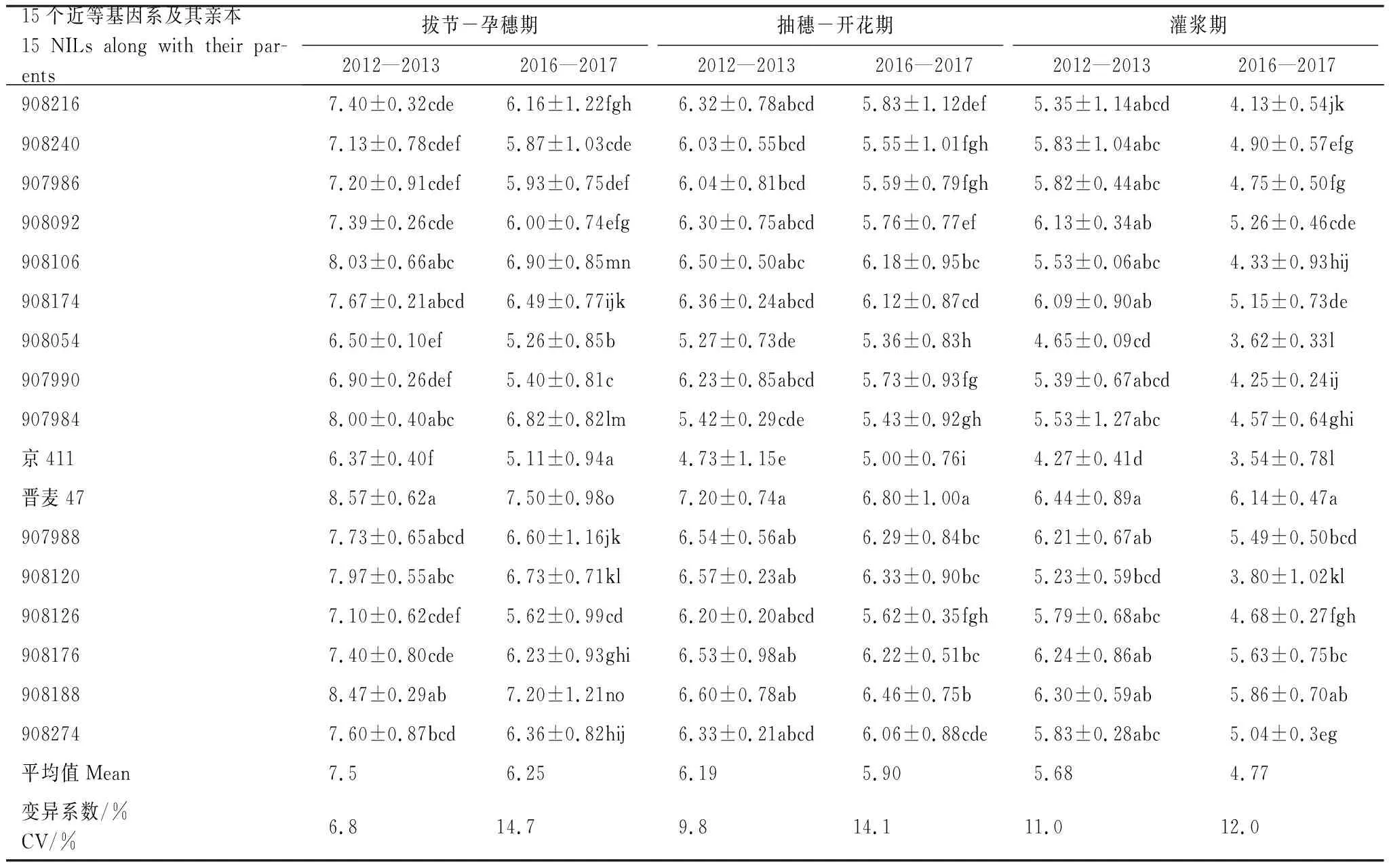
表1 冬小麦近等基因系及其亲本的冠气温差/℃
同一列标记不同字母的数值表示在P<0.05水平上有显著差异
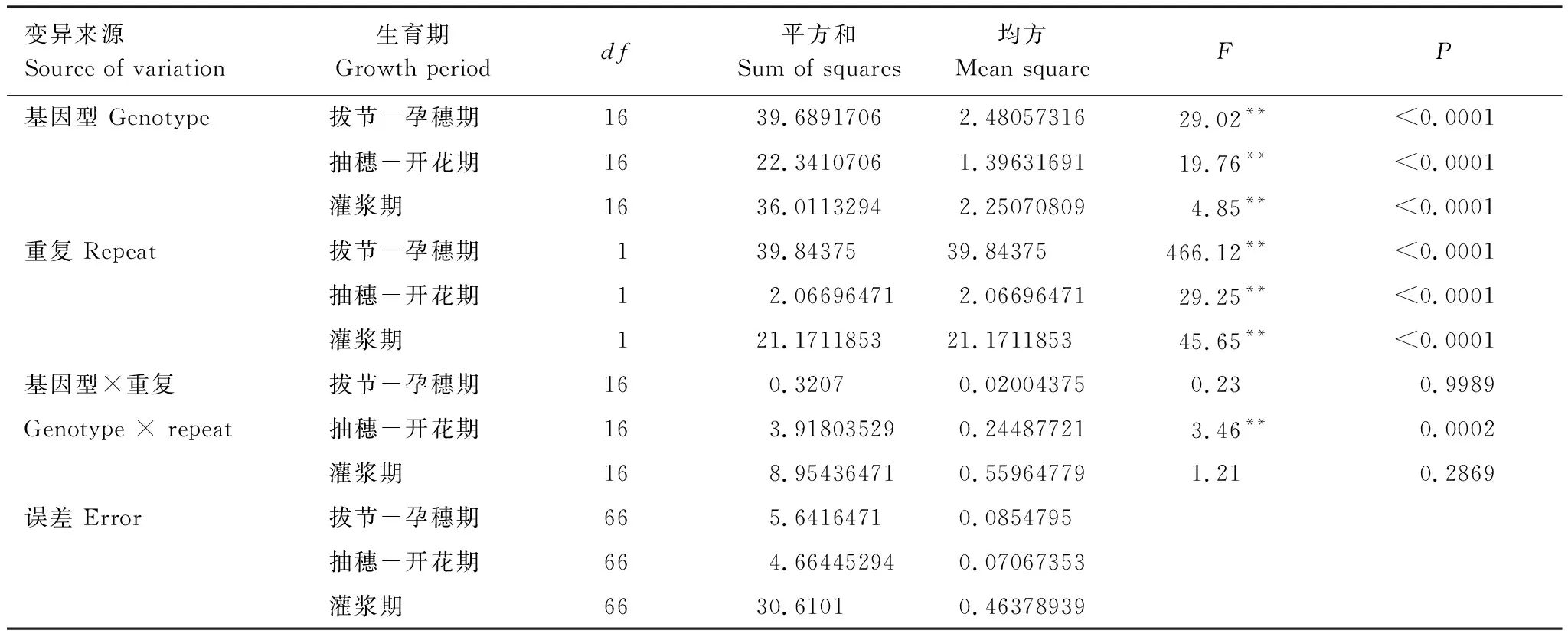
表2 冠气温差的方差分析
**在P<0.01水平上有极显著差异
与耗水量的情况不同,15个近等基因系及其亲本之间产量没有显著差异。主要源于耗水量的差异,品种/系之间WUE差异较大。2012—2013年度,参试冬小麦品系WUE分布在1.449—1.82 kg/m3范围内,平均值为1.64 kg/m3。907986最低,908274最高。亲本京411和晋麦47分别为1.52 kg/m3和1.68 kg/m3,二者之间达到显著差异。2016—2017年度,参试冬小麦品系WUE分布在1.554—1.857 kg/m3范围内,平均值为1.712 kg/m3。907986最低,908274最高。亲本京411和晋麦47分别为1.593 kg/m3和1.742 kg/m3,二者之间达到显著差异。
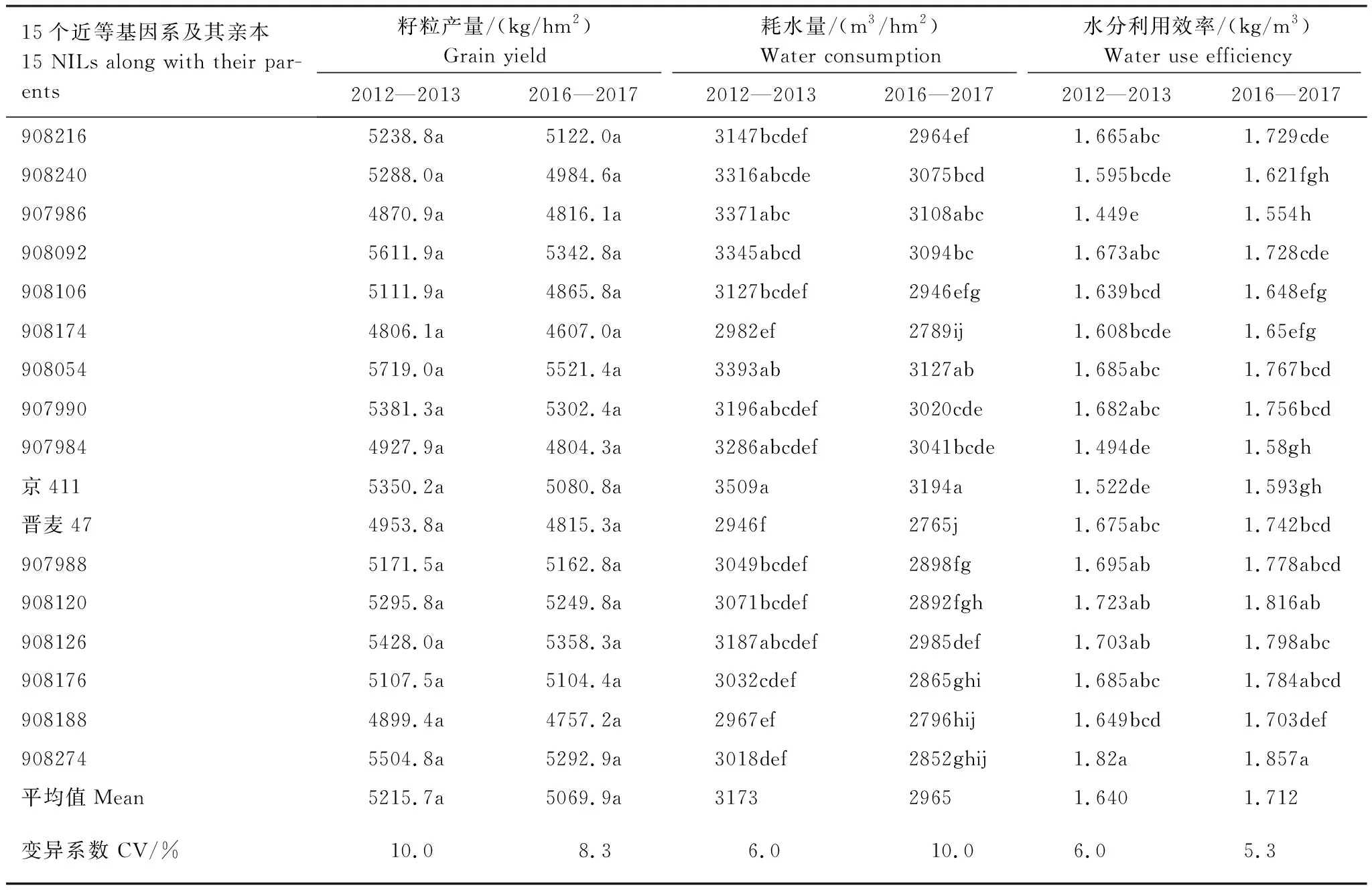
表3 冬小麦近等基因系及其亲本的产量、耗水量与水分利用效率
2.4 不同近等基因系及亲本的CTD与群体总耗水量的关系
拔节-孕穗期、抽穗-开花期和灌浆期测定的CTD与群体总耗水量均呈极显著负相关关系(图2),随着CTD增大,群体总耗水量减小。即在特定的生育期测定的CTD大的品种,群体总耗水量小;而CTD小的品种,群体总耗水量大。这种负相关关系在几个极端的品种/系中更加明显,相关性最强的抽穗-开花期,从两个年度的实验结果均可以看出,CTD最小的材料京411和908054,总耗水量均在最高一组,而CTD最高的材料晋麦47和908188,总耗水量均在最低一组。
CTD与总耗水量的相关性在拔节-孕穗期和抽穗-开花期相对较高,而在灌浆期则明显下降,2012—2013年度R2为0.4556,2016—2017年度R2为0.5424但是依然达到显著水平。R2最高峰出现在抽穗-开花期,两个年度分别为0.7042和0.6095,均达到极显著水平。从这两个年度的实验结果看,CTD每升高0.1℃,总耗水量分别下降近25 m3/hm2和29 m3/hm2。
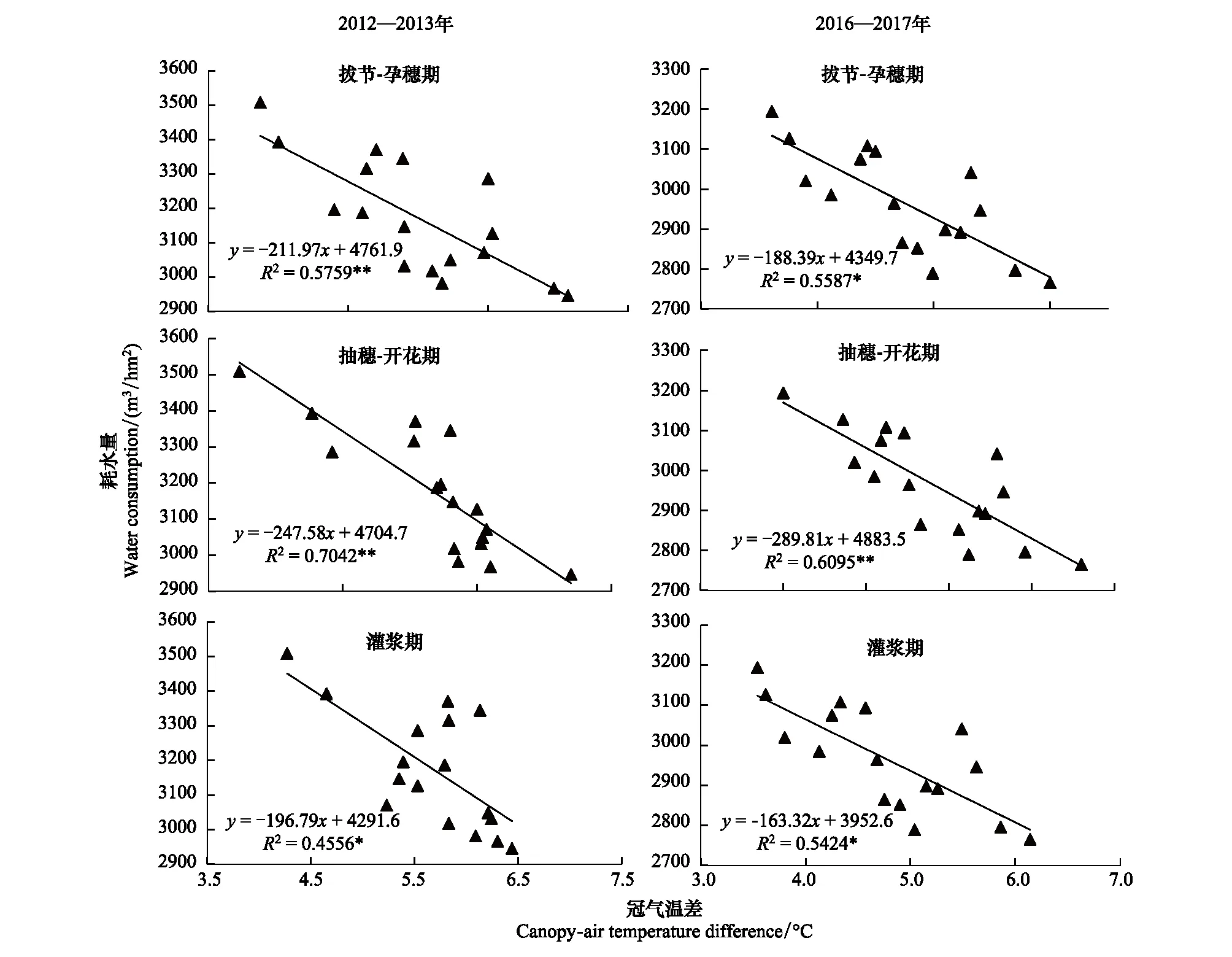
图2 冬小麦冠气温差与耗水量的关系 Fig.2 The relationships between CTD and amount of water consumed
2.5 不同近等基因系与亲本的叶片蒸腾速率和气孔导度与群体总耗水量的关系
2012—2013年度和2016—2017年度的实验结果均表明,叶片气孔导度度和蒸腾速率与群体总耗水量之间相关性很弱,两个年度均是拔节-孕穗期最高,R2分别为0.1477和0.2583,3个生育期均未达到显著水平(图3、4)。由此看来,如同干旱条件下叶片水平的光合速率与群体产量的相关性较小一样,叶片水平的蒸腾速率和气孔导度与群体总耗水量也不存在显著的相关性。这一结果表明,在水分亏缺条件下,该组近等基因系材料,叶片蒸腾速率和气孔导度不是影响群体总耗水量的关键因素。
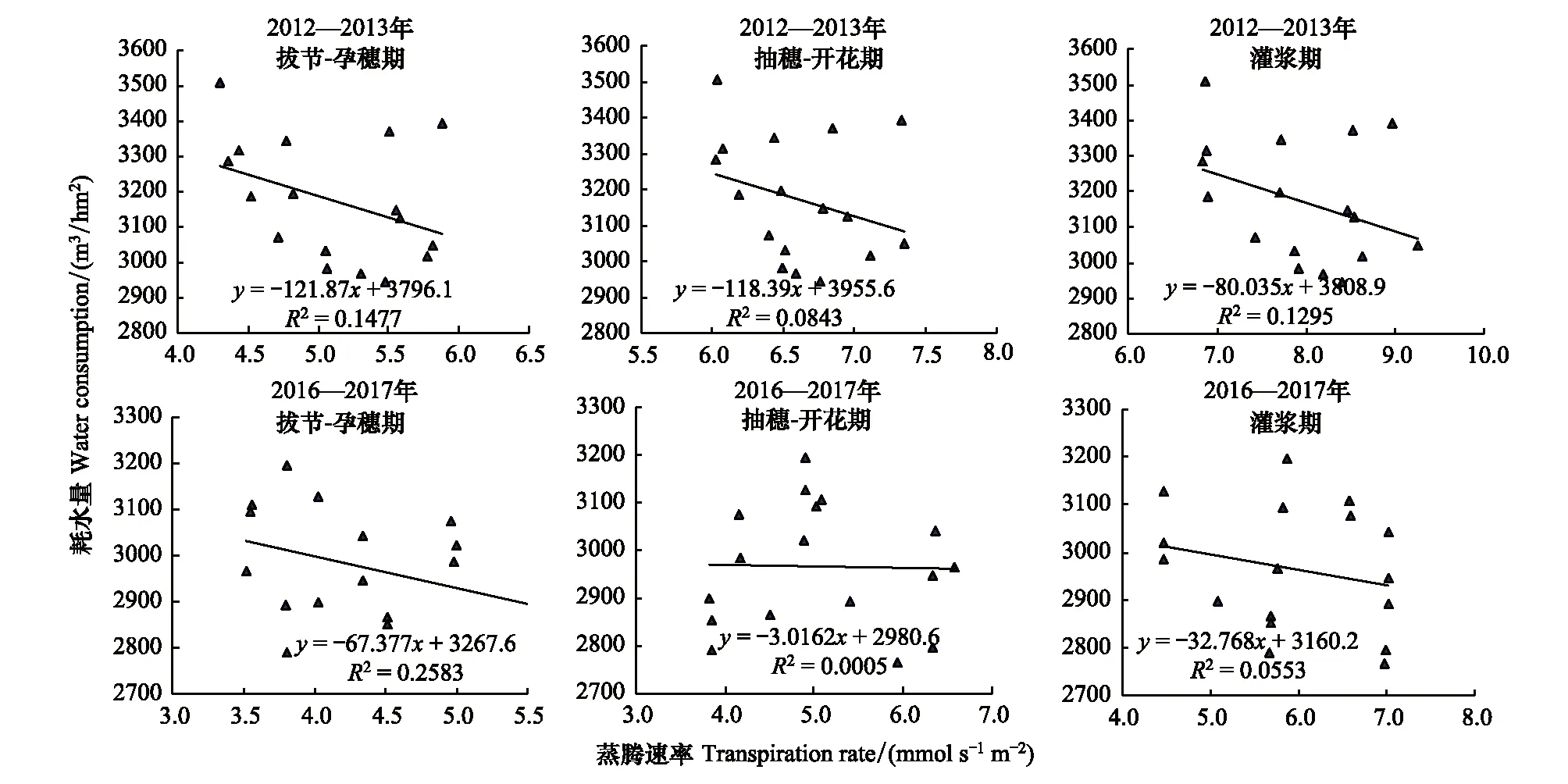
图3 冬小麦叶片蒸腾速率与总耗水量的关系Fig.3 The relationships between leaf transpiration rate and amount of water consumed
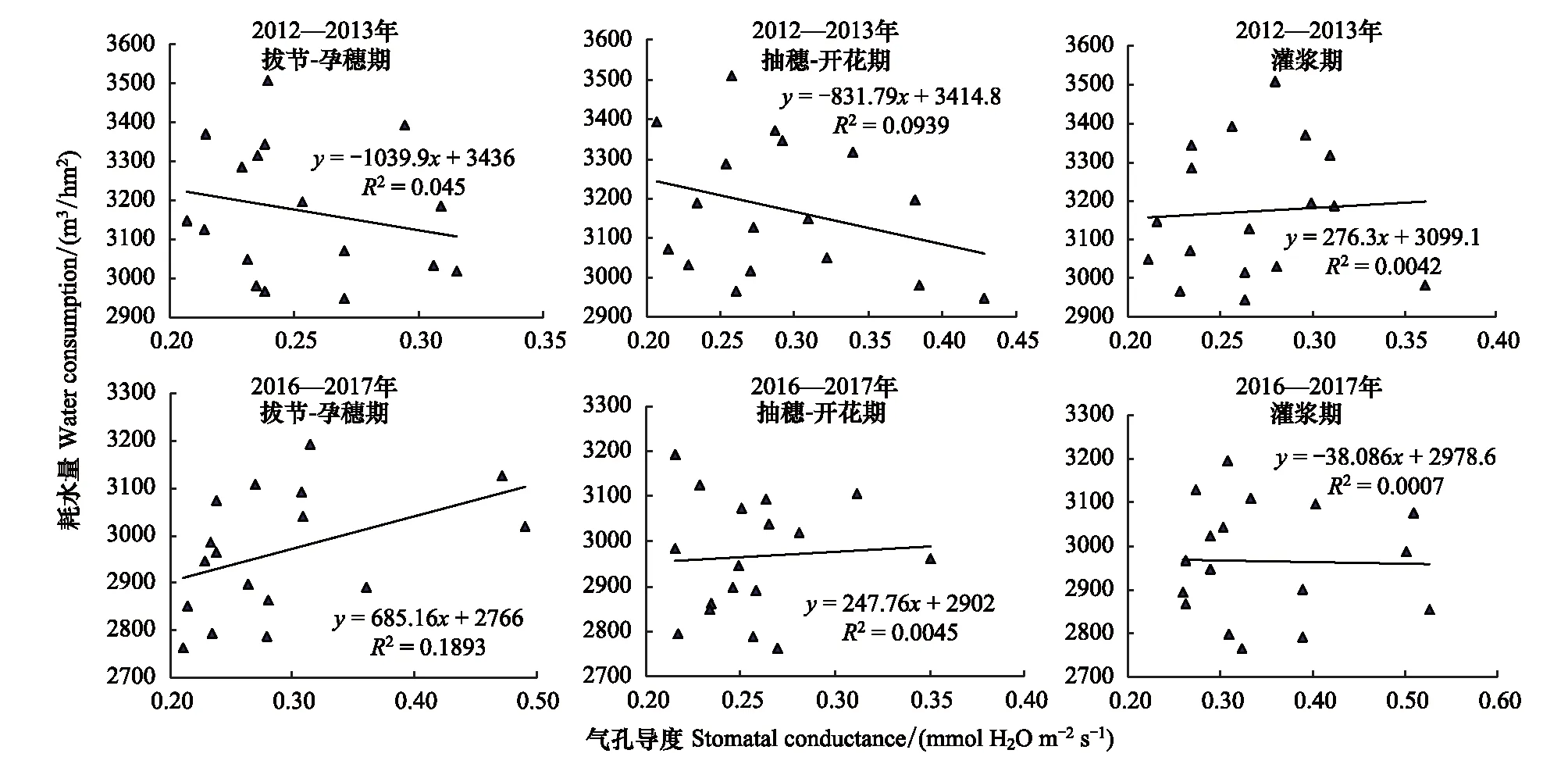
图4 冬小麦叶片气孔导度与总耗水量的关系Fig.4 The relationships between leaf stomatal conductance and amount of water consumed
3 讨论
一直以来,相同水分条件下品种间产量差异及其影响因素的研究备受关注,而产量相近的品种间耗水量的差异与其影响因素的研究却鲜有报道,而这正是探索节水品种鉴选方法的另一条重要途径。群体总耗水量受大气中饱和水汽压差、温度、风速、日照等气象因子、土壤水分条件、植物蒸腾特性及冠层结构特性等多种因素的影响[7-8,10]。在气象因素、土壤水分条件与种植密度相同的情况下,品种之间总耗水量差异的关键影响因素研究显然应该从品种自身生理特性与群体结构特性方面着手。本研究选用以晋麦47与京411为亲本的一组遗传背景相近的近等基因系为材料。其特点是产量差异不显著而群体总耗水量和CTD差异显著,这使我们得以从众多因子中单独分离出CTD,研究其对群体总耗水量的影响。结果首次揭示出CTD对群体总耗水量有重要影响,二者之间存在极显著负相关关系。另一方面,叶片蒸腾速率和气孔导度两项与耗水特性相关的重要生理指标与群体总水量相关性并不显著。作物冠层温度是由土壤-植物-大气连续体内的热量和水汽通量所决定,CTD是群体冠层结构特性相关的重要指标。本研究证明构建合理的群体冠层结构不仅是获得高产的途经,而且是调控群体总耗水量,提高品种水分利用效率的重要途径。
CTD与作物水分关系的研究主要集中于CTD作为土壤水分亏缺诊断指标方面[22-24]。而相同的土壤水分与气象条件下,不同品种之间CTD与耗水量关系的研究迄今尚无报道。强小曼等[25]和周颖等[26]探讨不同水分条件下CTD与蒸腾速率的日变化规律,发现二者变化基本一致。然而,这只是针对同一个品种随气象因子日变化CTD与蒸腾速率之间关系的研究,二者的主要限制因子为气象因子。与此不同的是,本研究是在相同的土壤水分与气象条件下,揭示品种之间耗水量差异及其与CTD之间的关系。由于环境条件一致,导致品种间耗水差异的主要因素则归因于作物群体结构特性与蒸腾生理特性[9,27-30]。品种间CTD的差异主要决定于群体冠层对水汽与热量的通透性。CTD与总耗水量之间极显著的负相关性表明,15个近等基因系及其亲本之间总耗水量的差异主要源于冠层结构特性的不同。依据前人的研究结果,群体内部植株间距、高度、及叶茎形状、数量与颜色等形态学特征构成特定的冠层结构[31-34],影响群体的荫蔽性与通风性,决定群体与大气之间水热交换的冠层阻力。冠层结构通透性强,冠层阻力小,土壤蒸发与群体蒸腾量大,即总耗水量大[5]。而群体与环境之间热量交换快,冠层内部及其与周围大气之间温度分异小[35],即CTD变小。据此,相同的环境条件与栽培措施下,CTD 大的品种,群体总耗水量会小。本研究结果即证实二者之间的极显著负相关关系。
不同生育期测定的叶片蒸腾速率和气孔导度与总耗水量的相关性均未达到显著水平。群体总耗水量的影响因素复杂,它不仅包括群体蒸腾耗水,还包括土壤蒸发耗水,既受环境因素的影响,又受品种蒸腾特性和群体冠层特性的制约。而且,群体内部的个体会通过强大的自调节能力进行群体适应。不同品种叶片蒸腾生理特性对单株耗水量或者群体在特定生育时期的耗水量起主要影响[1],而对整个生育期内总耗水量的影响可能被其他关键因素或者个体自调节作用抵消。本研究中CTD与总耗水量呈极显著负相关性,暗示品种蒸腾生理特性对群体总耗水量的影响可能被群体冠层特性的影响抵消,冠层特性为群体总耗水量的关键影响因素。
4 结论
在土壤水分亏缺程度相对一致的条件下,CTD差异显著的亲本品种晋麦47与京411及其15份近等基因系材料之间,群体总耗水量、不同生育期的CTD均存在极显著差异。群体总耗水量与拔节-孕穗期、抽穗-开花期和灌浆期测定的CTD之间均存在显著的负相关关系,R2在抽穗-开花期最高,两个试验年份分别达0.7042和0.6095。而群体总耗水量与叶片蒸腾速率和气孔导度之间相关性极弱,两个试验年份3个生育期测定的结果均未达到显著水平。这些结果表明,水分亏缺条件下,该组近等基因系材料中,影响群体总耗水量的关键因素不是叶片蒸腾生理特性,而是群体冠层结构特性。构建合理的群体冠层结构是调控群体耗水量,提高品种水分利用效率的重要途径。
致谢:中国农业科学院作物科学研究所景蕊莲研究员为本研究提供近等基因系材料,并对研究方案与实验设计给予帮助,特此致谢。
猜你喜欢
绿色科技(2022年7期)2022-11-16
中国农业信息(2022年1期)2022-05-25
农业机械学报(2021年11期)2021-12-07
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
农业科技与信息(2021年8期)2021-05-17
北方果树(2020年6期)2020-11-14
中国农业科技导报(2020年12期)2020-03-15
农业环境科学学报(2017年2期)2017-03-20
Advances in Polar Science(2015年1期)2015-02-06
