
来源地纬度和分解者类群对中国东部滨海湿地芦苇叶凋落物分解速率的影响
2019-02-10 03:04刘子夏张有政吴纪华
复旦学报(自然科学版) 2019年6期
刘子夏,张有政,吴纪华
(复旦大学 生命科学学院 生物多样性和生态工程教育部重点实验室,上海 200438)
植物凋落物是全球陆地碳库的重要组成部分,其总量高达300PgC[1-2],约占全球陆地表层总碳储量的1/7[3].植物凋落物通过分解将土壤表层碳转移到大气碳库中,是全球陆地碳通量的主要来源[4],每年产生的碳通量近58PgC,约占全球陆地总碳通量的一半[2,5],因此植物凋落物分解速率的变化会直接导致土壤和大气碳库的变化.盐沼是陆地上碳沉积速率最快、固碳能力最强的生态系统之一[6],盐沼生态系统中土壤碳库对于盐沼植物凋落物分解速率变化的响应比其他陆地生态系统更加敏感.因此,研究盐沼植物凋落物的分解作用对了解盐沼生态系统碳循环过程有着重要的作用.
植物凋落物分解速率的变化受到诸多因素的调控.已有的研究报道发现,气候、凋落物质量和分解者类群均对凋落物分解速率有重要影响,这些影响因素在不同植物种类和实验环境下影响的显著性和程度不同,各个影响因素之间是否有交互作用也存在争议.早期的研究提出凋落物分解速率同时受到气候和凋落物质量的影响[7-8],这一结论在大量相关研究中被印证.如Berg等对欧洲和美国东部针叶林凋落物分解速率的影响因素进行了研究,发现分解地年实际蒸腾量对凋落物分解速率的总方差解释率达到49.6%,同时凋落物中氮、磷和水溶性物质的含量对同一分解地的凋落物分解速率方差解释率大于90%[9].Zhou等对全国12个地区的植物凋落物进行原位分解实验,发现植物凋落物分解速率受气温、降雨量、凋落物碳氮比和氮磷比的调控[10].而在Irons等的研究中则发现美国阿拉斯加和密歇根地区的木本植物叶凋落物分解速率无显著差异,因此认为气温这一气候因素在高纬度地区并不是影响凋落物分解速率的主要因素,而不同地区的分解者可能对凋落物分解速率有重要贡献[11].分解者作为凋落物分解过程的直接参与者,同时也受到诸多环境因素的控制,Garcia-Palacios等通过整合分析包含热带雨林、落叶林、农田、干旱或寒冷地区以及草甸生态系统的440个植物凋落物分解实验结果发现,土壤动物作为分解者对于凋落物分解速率的影响在全球和地区研究尺度上均显著,在土壤动物存在的条件下分解速率平均提高27%;但土壤动物对于凋落物分解速率的影响程度受气候和凋落物质量共同调控,且在不同生态系统中调控程度不同[12].
纬度梯度格局是生态学研究的重要议题之一,它综合反映了温度、降水和光照等因子在纬度上的梯度变化.已有研究表明新鲜植物叶片的质量在不同纬度和生长条件下存在差异,在针对全球新鲜植物叶片质量的研究中,发现全球尺度下植物叶片中氮、磷和氮磷比均与气温呈正相关关系,与纬度呈负相关关系[13].而植物新鲜叶片质量差异所导致的叶片凋落物质量的差异对于植物凋落物分解速率存在潜在的影响,同时也可能会进一步与分解者产生交互作用从而影响凋落物分解速率.在森林和草甸生态系统中,纬度梯度会导致植物凋落物可分解性的差异,进而形成区域碳动态的纬度差异格局[14].目前在盐沼生态系统中对于植物凋落物分解速率的跨区域尺度研究较少,涉及盐沼植物凋落物可分解性纬度格局的研究未见报道,由来源地纬度导致的凋落物可分解性差异与分解者类群对凋落物分解速率的影响是否存在交互效应也尚不清楚.
芦苇(Phragmitesaustralis)作为全球盐沼生态系统的广布种,在我国盐沼生态系统中大量分布.在芦苇占优的盐沼生态系统中,芦苇凋落物分解的速率差异直接导致了生态系统碳库的变化.本研究以来自中国沿海9个盐沼生态系统的芦苇叶凋落物为研究对象,在上海长江口湿地进行同质园凋落物分解实验,试图探究: (1) 中国盐沼生态系统中芦苇叶凋落物可分解性的纬度格局;(2) 分解者类群、来源地纬度及两者交互效应对芦苇叶凋落物分解速率的影响.
1 材料与方法
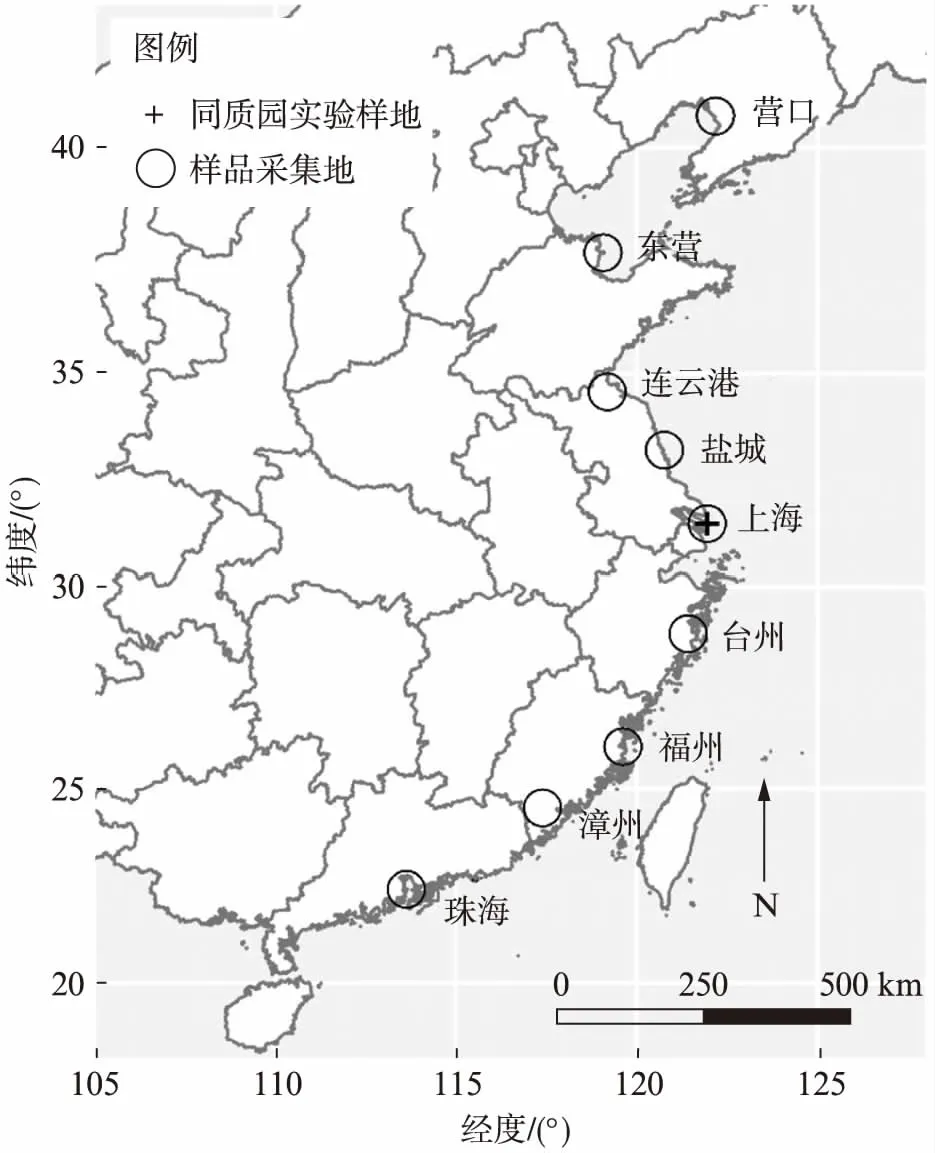
图1 样品采集地及同质园实验样地空间分布图Fig.1 Map of sample collection sites and common garden experiment site
1.1 样品的采集和处理
本实验于2015年8—11月份采集了中国东部沿海9个地区的盐沼芦苇叶凋落物,南至珠海北至营口(图1).在每个地区,间隔1km以上进行5组重复采样(每组选取5m×5m的样方,在里面随机收取5个1m×1m的小样方叶凋落物,然后混合),共计获得45组芦苇叶凋落物样品.凋落物样品选取依据参考廖成章等[15]的方法,选取颜色鲜黄、韧性强、茎秆上有枯叶相连的新叶凋落物,分装后带回实验室清洗去除叶凋落物表面的污泥后放入60℃烘箱中烘干至恒重.
对45组叶凋落物样品分别测定物理指标: 叶厚度、叶韧性、比叶面积(叶面积/叶质量)和化学指标: 碳、氮、磷、钾、钙、镁、单宁、木质素、纤维素、半纤维素、总营养元素含量、总酚含量,其中化学指标参考国家标准、农业标准和ISO国际标准方法(NY/T 2017—2011、GB/T 35818—2018、ISO 14502—1: 2005).

表1 凋落物袋孔径对应的分解者类群
洗净烘干的45组叶凋落物样品被分装到5mm、2mm、0.1mm孔径的3种凋落物袋中,凋落物袋孔径的选择参考了Bradford等对凋落物袋孔径和土壤动物类群的对应关系(见表1)的描述[16].每个凋落物袋中称取(15±0.5) g叶凋落物样品,每组叶凋落物×凋落物袋孔径组合重复3次,共计获得3份135个凋落物袋实验单元共计405个.对每个凋落物袋单元进行标记并记录下分解前的凋落物净重量.
1.2 凋落物同质园分解实验
同质园实验样地位于上海市长江口北湖区域(东经121.65°,北纬31.68°,图1),该区域为以芦苇为优势种的自然盐沼生态系统,受到人为扰动极少.在同质园实验样地中设置3个实验重复样点,每个样点之间间隔超过1km以避免假重复现象.3个样点的土壤本底信息见表2.
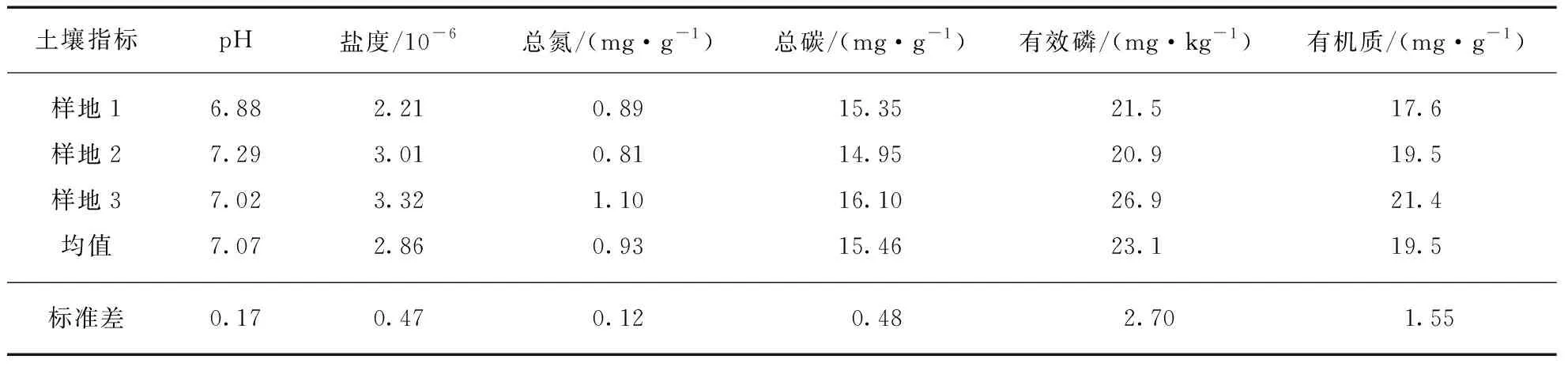
表2 分解实验样点土壤理化信息
于2017年8月4日将3份凋落物袋实验单元分别埋入3个样点的10cm深土层中(凋落物袋埋藏法,参考TBI标准[17]),同样点内的凋落物袋实验单元随机排布,盖上土后待其自然分解.3个月后,于2017年11月3日将凋落物袋实验单元挖出后分别装袋带回.将凋落物袋单元放在150目(0.1mm孔径)的网筛中进行清洗,洗去污泥并同时挑除进入凋落物袋中的植物根系.清洗后放入60℃烘箱中烘干至恒重,称重记录下分解后的凋落物净重量.
1.3 计算与分析
在本研究中,我们使用Olson负指数衰减模型[18]计算凋落物分解速率常数k,这一方法广泛使用于凋落物分解速率研究用于比较分解速率的快慢,k值越大代表分解速率越快,其计算公式为:
其中:k为分解速率常数(a-1),M0为分解前的凋落物质量,Mt为分解后的凋落物剩余质量,t为分解时间(以年为单位).
在实验过程中有21个样本丢失或凋落物袋破损严重,因此使用平均值法插入该组处理(45种凋落物×3种凋落物袋孔径组合相同的为一组)的平均值来代替缺失的数据.
使用Bartlett方差齐性检验(Bartlett test)检查确定样本满足方差分析要求,使用二元方差分析(2way-ANOVA)比较不同来源地和不同凋落物袋孔径对凋落物分解速率的影响,并且使用Tukey事后检验(TukeyHSD test)比较各处理组之间的凋落物分解速率差异显著性,以95%置信区间是否重合作为检验标准.凋落物可分解性纬度格局和凋落物质量指标纬度格局使用线性回归(linear regression)进行拟合,用于分别检验凋落物可分解性和凋落物质量指标与凋落物来源地纬度的相关性.为了在同一量纲水平上比较凋落物质量指标,使用标准分计算法转换得出对应的凋落物质量指标相对值,其计算公式为:
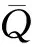
本实验通过凋落物袋孔径来控制凋落物所暴露的分解者类群,但由于凋落物袋孔径由小至大其所暴露的分解者类群是依次叠加的,因此如果要比较每个分解者类群对凋落物分解的贡献率则需要进行额外的计算,其计算公式为:
其中:k0.1、k2、k5分别代表凋落物袋孔径为0.1mm、2mm和5mm条件下的凋落物分解速率常数k的均值,E2、E5分别代表中型和大型土壤动物对凋落物分解的净贡献率.
本研究中的统计分析、图表绘制均使用R语言版本3.4.3(R Core Team 2017)进行,其中方差分析使用基础函数库中aov函数,Tukey事后检验使用基础函数库中TukeyHSD函数,线性回归模型使用基础函数库中lm函数.图表绘制使用扩展包: agricolae、ggplot2、ggpmisc、maptools、multcomp.
2 结 果
2.1 芦苇叶凋落物的分解速率与其来源地纬度的关系
不同来源地的芦苇叶凋落物分解速率存在显著差异(表3).由来源地不同所导致的芦苇叶凋落物分解速率差异的均方为15.872,占芦苇凋落物分解速率总体方差的72.16%,说明芦苇叶凋落物的来源地是导致分解速率差异的主要因素.由芦苇叶凋落物分解速率与来源地纬度的线性回归(图2)结果可知: 芦苇叶凋落物在3种凋落物袋孔径下的分解速率均与来源地纬度呈极显著正相关关系,即来源地纬度越高的芦苇叶凋落物分解速率越快.

表3 芦苇叶凋落物来源地和凋落物袋孔径对其分解速率的影响

图2 不同凋落物袋孔径下芦苇叶凋落物分解速率与来源地纬度的关系Fig.2 Relationship between decomposition rate of Phragmites australis leaf litter and latitude of source area under different mesh sizes of litter bag
2.2 芦苇叶凋落物分解速率与其所暴露的分解者类群的关系
不同凋落物袋孔径下的芦苇叶凋落物分解速率存在显著差异,且与凋落物来源地纬度无显著交互作用(表3).由凋落物袋孔径不同所导致的芦苇叶凋落物分解速率差异的均方为4.85,占芦苇凋落物分解速率总体方差的22%,说明分解者类群也是芦苇叶凋落物分解速率的影响因素,但其影响程度相比凋落物来源地而言较低.在凋落物孔径为0.1mm、2mm、5mm的条件下,芦苇叶凋落物分解速率与其来源地纬度的线性回归(图2)方程截距分别为0.6118、0.1843、0.3585,表明小型土壤动物及微生物对芦苇叶凋落物分解速率的贡献最大,而中型土壤动物对凋落物分解速率存在负效应.从分解速率均值中型和大型土壤动物对凋落物分解速率的贡献率(表4,见第682页)可以看出,中型土壤动物对于几乎所有来源地(除连云港外)的芦苇叶凋落物分解速率的贡献均为负,且对于来源地为盐城和东营两地的芦苇叶凋落物分解速率有显著的负效应;大型土壤动物对芦苇叶凋落物分解速率的贡献正负不一,仅对来源地为福州的芦苇叶凋落物分解速率有显著的负效应.
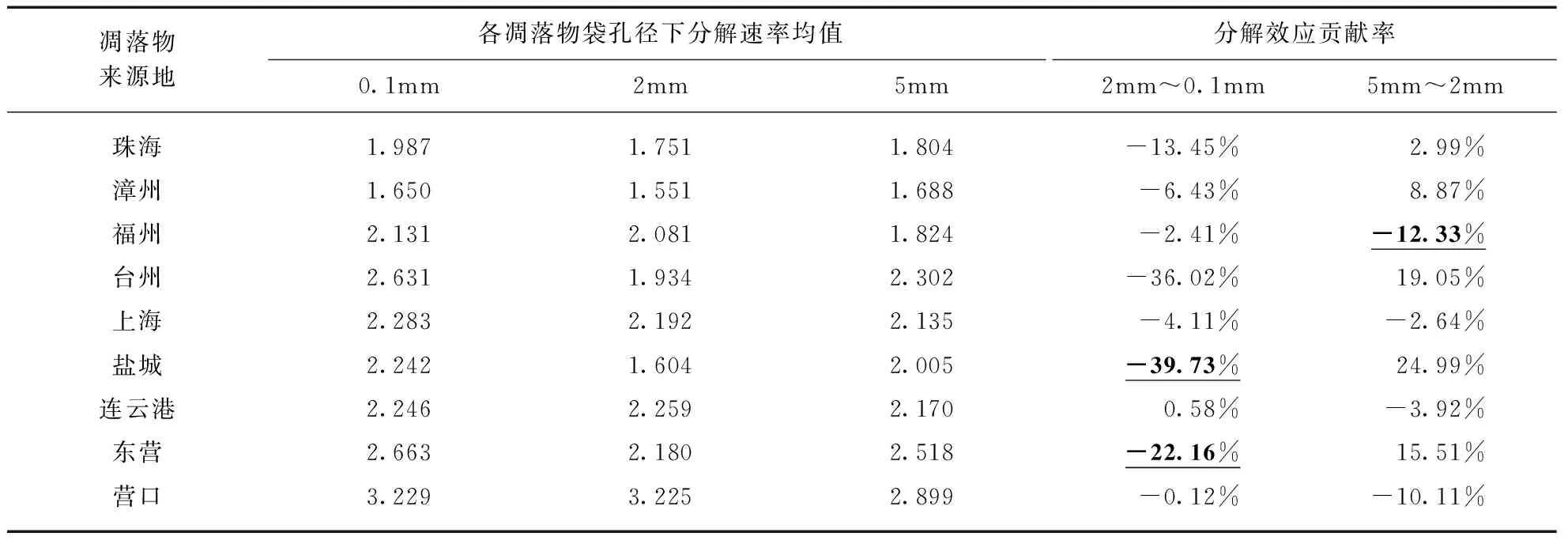
表4 不同分解者类群对芦苇叶凋落物分解的影响及贡献率
注: 下划线代表效应显著(基于TukeyHSD检验)
2.3 芦苇叶凋落物质量指标与来源地纬度的相关关系
不同来源地的芦苇叶凋落物质量存在差异.分析芦苇叶凋落物质量指标与其来源地纬度的线性回归结果见表5.由表5可以看出,所测芦苇叶凋落物质量指标中除酚磷比、叶韧性与来源地纬度的相关回归不显著外,其他叶凋落物质量指标均与其来源地纬度呈显著线性相关.其中纬度差异对凋落物质量差异解释率大于30%(即R2>30%)的凋落物质量指标有碳、氮、钙、总营养元素、总酚、纤维素含量,碳氮比、氮磷比、单宁氮比、纤维素氮比以及叶厚度,共计11个凋落物质量指标.

表5 芦苇叶凋落物质量指标与其来源地纬度的线性回归
3 讨 论
3.1 芦苇叶凋落物可分解性的纬度格局与其凋落物质量指标间的关系
植物凋落物的可分解性(plant litter decomposability)是衡量不同的植物凋落物样品在相同的分解条件下的分解速率快慢的相对指标,也是植物凋落物质量对分解速率影响的体现[19-20].本研究发现在相同的凋落物袋孔径下芦苇叶凋落物可分解性存在着随来源地纬度升高而增加的格局,这一格局的成因本质上是一系列芦苇叶凋落物质量指标的来源地纬度格局.通过对本实验中芦苇叶凋落物分解速率与其质量指标进行相关分析发现: 芦苇叶凋落物分解速率与叶凋落物的氮、总营养元素、钙、钾、镁、钠、磷含量以及叶韧性、叶厚度等凋落物质量指标呈正相关,与叶凋落物的碳、单宁、木质素、总酚、纤维素、半纤维素含量和碳氮比、碳磷比、氮磷比以及比叶面积等凋落物质量指标呈负相关(筛选出线性回归分析中P<0.05且R2>20%的凋落物质量指标绘制图3).

图3 芦苇叶凋落物分解速率与其质量指标相关关系Fig.3 Relationship between decomposition rate of Phragmites australis leaf litter and its quality index注: 图中回归检验的P值均小于0.001
在以往的研究结论中,凋落物的可分解性与凋落物中难分解组分含量呈负相关,与凋落物中易分解组分含量呈正相关.如Pérez-Harguindeguy等在对阿根廷木本植物叶凋落物分解速率的研究中发现: 叶凋落物分解速率与其氮含量呈正相关,与叶凋落物中碳含量、碳氮比呈负相关[21].Vaieretti等的研究结果同样支持了上述结论,同时还发现叶凋落物分解速率与木质素-纤维素-半纤维度指数(Lignin-Cellulose-Hemicellulose Index/LCH Index)呈负相关[22].本研究结果验证了上述结论: 半纤维素作为难分解组分,其在芦苇叶凋落物中的含量与芦苇叶凋落物分解速率呈极显著负相关,且方差解释率达到40%,说明芦苇叶凋落物中半纤维素是其分解速率的重要控制因素.
除了凋落物中难、易分解组分占比对凋落物分解速率有显著影响外,凋落物的营养组分和离子含量以及物理特性对凋落物分解速率也有显著影响.Trofymow等在研究加拿大18个森林生态系统的叶凋落物分解速率时,将氮、磷、硫、钾、钙、镁作为凋落物特性指标进行模型拟合,发现用凋落物的氮、磷、钙含量可以很好的解释凋落物分解速率的差异[23].Zhang等分析了70篇研究论文中110个区域的凋落物质量分解速率数据发现,凋落物分解速率与其磷、钾、钙、镁含量呈正相关[24].本研究也发现,芦苇叶凋落物的钙、镁、钠等金属离子以及磷和总营养元素含量与芦苇叶凋落物分解速率呈极显著负相关,且方差解释率均在20%以上.另外芦苇叶凋落物中碳、单宁、总酚和纤维素与磷的比值与芦苇叶凋落物分解速率呈显著负相关.表明金属离子和营养元素对芦苇叶凋落物分解速率也有重要影响.
有研究表明凋落物的物理特性与凋落物分解速率也存在相关关系.Zukswert等发现凋落物分解速率与比叶面积呈正相关与叶厚度呈负相关[25],Pérez-Harguindeguy等的结果中同样也提到了凋落物分解速率与叶韧性呈负相关[21].但本研究中芦苇叶凋落物的物理质量指标与凋落物分解速率的相关关系与上述结论相反: 芦苇叶凋落物的叶韧性越强分解越快、比叶面积越大分解越慢.单独分析叶凋落物物理质量指标与分解速率的关系,上述结果无法找到合理的解释,但是由于凋落物质量指标之间存在共线性,因此芦苇叶凋落物物理质量对凋落物分解速率的影响可能被化学质量指标所掩盖,从而导致出现于预期相反的结果.
3.2 分解者类群对凋落物分解速率的影响
目前关于分解者类群对植物凋落物分解速率影响相对重要性的研究结果存在较大争议,不同分解者类群对植物凋落物分解速率的影响程度也因生态系统不同而存在差别.在多数分解者对凋落物分解速率的影响研究中,大中小型土壤动物和土壤微生物均对凋落物分解速率有显著影响,即凋落物袋孔径越大凋落物分解速率越快[16,26-27].但是本实验结果表明: 对于来源地为福州、盐城和东营的凋落物样品在2mm孔径凋落物袋中的分解速率显著低于在0.1mm孔径的凋落物袋中的样品,即对应的中型土壤动物对凋落物分解速率存在负效应,而其他来源地的凋落物在不同凋落物袋孔径下分解速率无显著差异(表5).导致这一结果的原因可能来自于两个方面: 1. 盐沼生态系统中微生物和小型土壤动物在凋落物分解过程中占主导,在Yang等对中国东海岸土壤动物和叶凋落物分解速率的研究中发现,小型土壤动物对凋落物分解的贡献是大型土壤动物的6倍[28];2. 中型土壤动物中食微生物土壤动物的存在抑制了微生物的数量和活动,在Hanlon等的研究发现中型土壤动物对微生物的捕食效应显著抑制了微生物的呼吸强度[29].
在凋落物分解研究中使用凋落物袋孔径控制分解者类群是惯用的方法,最早由Bocock等提出并使用[30],但是仅通过凋落物袋孔径控制所暴露的分解者类群存在诸多缺陷.如上文所提到的中型土壤动物对微生物的捕食效应,在较大凋落物袋孔径下,凋落物不仅暴露在了中型、大型土壤动物分解者下,同时微生物、小型土壤动物分解者也暴露在了中型、大型土壤动物捕食者下,导致原有的分解效应被抑制.另外即使是大孔径的凋落物袋也会阻止大型无脊椎动物参与凋落物分解,特别是在盐沼生态系统中有重要食碎屑作用的螃蟹等[31].因此要更加准确地揭示不同分解者类群对凋落物分解的贡献需要使用更加精确的控制手段.Patoine等在实验中使用灭菌的土壤,通过添加不同的优势土壤动物分解者,包括4种小型节肢动物和一种环节动物进行凋落物分解实验,比较了暴露在不同分解者类群下的凋落物分解速率[32],这一方法为研究凋落物分解过程中分解者类群对分解速率的贡献提供了更精确的途径.
4 结 论
芦苇叶凋落物分解速率受凋落物来源地纬度和分解者类群的共同作用,其中凋落物来源地纬度的影响占主导,二者之间无显著交互效应.芦苇叶凋落物可分解性与纬度呈显著正相关关系,凋落物来源地纬度越高越容易分解,这一格局可以被芦苇叶凋落物磷、钙、钠、镁、总营养元素、半纤维素的含量和其碳磷比等凋落物质量指标的纬度格局很好地解释.本文首次通过同质园实验得出了我国东部沿海跨区域尺度的芦苇叶凋落物可分解性纬度格局.这一格局表明,仅通过净初级生产力、和凋落物碳总量来计算盐沼生态系统碳库及碳通量是不够的,还需要考虑到不同纬度地区凋落物易分解的差异可能导致的估算偏差.因此本文的研究结果为大尺度盐沼生态系统碳库和碳循环估算提供了新的思路.本研究以同质园实验揭示了来源地不同的芦苇叶凋落物可分解性差异,但是在实际条件下,芦苇凋落物是在当地产生和分解的,而单一地点实验没有考虑分解地环境导致的凋落物分解速率变化,因此需要进行跨区域尺度的交互移植实验探究气候、分解者和凋落物质量共同影响下的凋落物分解速率变化.
猜你喜欢
人民黄河(2020年12期)2020-12-30
山西农业科学(2020年9期)2020-09-14
——乌尤尼盐沼
家教世界(2020年17期)2020-07-06
山西农业科学(2019年12期)2019-12-19
环球时报(2019-11-14)2019-11-14
测绘学报(2019年4期)2019-06-10
百科探秘·航空航天(2017年10期)2017-11-08
生态与农村环境学报(2017年8期)2017-08-22
齐鲁周刊(2017年29期)2017-08-08
电子制作(2017年10期)2017-04-18
