
同域分布3种啄木鸟冬季取食的生态位差异
2019-01-18 05:36:10司雨蕙潘麒嫣
生态学报 2018年23期
戎 可, 司雨蕙, 潘麒嫣, 王 欢
1 东北林业大学野生动物资源学院, 哈尔滨 150040 2 国家林业局野生动物保护学重点开放实验室,哈尔滨 150040
生物多样性的形成与维持无疑是生态学最具吸引力的研究领域之一[1]。探讨物种如何共存,更有助于提高自然保护工作的有效性[2]。生态位分化假说(niche differentiation)部分解释了物种共存的机制[3- 4],也受到很多研究的支持[1,5- 6],但有关同域分布啄木鸟生态位差异的研究却很少[7]。
现存啄木鸟科(Picidae)动物有254种,分布于除大洋洲和南极洲外的其他大陆[8]。大部分啄木鸟会凿木取食蛀干昆虫,因此常被认为是控制森林害虫最具潜力的生物防治者[9]。更为重要的是,作为初级凿洞者(primary cavity excavator),啄木鸟在树干上挖掘的树洞,为其他动物的繁殖和栖息提供了生态位[10]。于是,不同的洞栖动物间形成了复杂的物种间的巢洞利用网(nest web)[11],成为提高和维持森林生物多样性水平的基础。啄木鸟也因此成为森林生态系统健康的指示物种和森林生态系统保护的伞护种[12-13]。所以,研究啄木鸟的生态学特征,对于理解森林生态系统的演替规律,保护森林生态系统具有重要意义,然而相关的研究还很缺乏[14],在国内更是寥寥无几[15-16]。
我国现有29种啄木鸟,其中10种分布于东北地区[17]。在小兴安岭的阔叶红松林中,黑啄木鸟(Dryocopusmartius)、三趾啄木鸟(Picoidestridactylus)和大斑啄木鸟(Dendrocoposmajor)是较为常见的3种啄木鸟,同时又都是靠捕食蛀干昆虫越冬的鸟种[18-20]。这些啄木鸟如何共存于食物资源有限的环境,是值得研究的主题[15-16]。本文将以生态位分异假说出发,报道这3种啄木鸟在冬季食物资源利用上的差异。
1 材料与方法
1.1 研究地概况
本研究的野外工作在黑龙江省凉水国家级自然保护区(以下简称凉水保护区)核心区开展。凉水保护区的主要保护对象是以红松为主的温带针阔混交林生态系统,属森林和陆生野生动物类型的自然保护区。保护区内森林类型包括原始阔叶红松林、原始冷云杉林、次生白桦林、次生山杨林、次生白桦山杨林、次生硬阔叶林、人工红松林、人工落叶松、人工云冷杉林和人工樟子松林等[21]。保护区现有鸟类16目46科252种,兽类6目16科44种[21]。其他概况见参考文献[22]。近年来,随着保护工作的加强,保护区境内生物多样性水平逐渐提高,原始林逐步向过熟林演替,啄木鸟变得愈发常见(待发表数据)。
1.2 数据收集方法
1.2.1 野外调查时间
采用样线法结合样方法进行野外数据收集,野外调查时间2016年1月5日至2016年1月13日。每天调查开始于6:30,结束于17:30。每天5组调查人员同时开展调查。
1.2.2 样线、取食样方及对照样方设置
在凉水保护区核心区内,地势最低的凉水河两侧,依东西走向分别均匀布设样线,样线间隔500 m,样线长1 km,共布设样线45条。每条样线各往返调查一次,因地势不同,往返耗时需2—4个小时。其中25条样线于上午完成调查,20条样线下午完成调查。
样线调查过程中,每遇啄木鸟取食,即观察并记录其取食行为特征,等啄木鸟离开取食树后,以取食树为中心设置取食样方,测量并记录取食生境特征。为了统计取食树周围树木密度,样方大小为20 m×20 m。为保证数据的独立性,往返同一条样线时,如在同一地遇到同种啄木鸟取食,则视为重复样方,不做观察和记录。调查中共获得取食样方312个。
本研究使用基于利用-可利用方法(use-available method)的Bailey′s方法[23]进行啄木鸟取食生境选择分析,因此需要设置对照样方。对照样方布设在调查样线上,每隔100 m设一个,实际共设置484个,对照样方大小与取食样方相同。
1.2.3 数据收集内容及方法
在取食样方中,除鸟种、性别、取食时间外,另收集14项取食生境因子及取食行为特征数据。具体野外记录数据包括(括号内为记录变量名):
(1) 鸟种(BIS),黑啄木鸟记为BW,三趾啄木鸟记为TT,大斑啄木鸟记为GS;
(2) 性别(SEX),啄木鸟的性别,雄性记为1、雌性记为2;
(3) 取食时刻(TM),以遇见时间记录至最近的0.5 h时刻;
(4) 林型(FOT),划分为天然林(记为1)、次生林(记为2)和人工林(记为3);
(5) 坡度(SLP),划分为坡顶(记为1)、中坡(记为2)和谷地(记为3);
(6) 优势树种(DTS),样方内优势树种,划分为冷杉(记为1)、云杉(记为2)、红松(记为3)和阔叶树种(记为4);
(7) 郁闭度(CRD),林冠层郁闭度,在样方四角,两个人分别目测后取平均数,精确到小数点后1位;
(8) 树密度(TRD),样方内胸径15 cm以上乔木的数量;
(9) 死树密度(SND),样方内胸径15 cm以上死树(包括倒木)的数量;
(10) 取食树种(FTS),取食时所在树种,调查时记录到具体树种,数据分析时划分为冷杉(记为1)、云杉(记为2)、红松(记为3)和阔叶树种(记为4);
(11) 取食树高(HET),啄木鸟取食时所在树的高度,依据三角测量法使用手持激光测距仪(Nikon COOLSHOT 40)测量,计算精度到0.5 m;
(12) 取食树胸径(DBH),啄木鸟取食时所在树胸径,使用胸径尺测量,精确到1 cm;
(13) 取食树状态(TSI),啄木鸟取食时所在树的存活状态,划分为活树(记为1)、死树(记为2);
(14) 取食高度(FHE),啄木鸟取食时所在的垂直位置,划分为冠层(记为1)、树干(记为2)、树基或倒木(记为3)。树基指树干基部2 m以下的部分;
(15) 取食部位(FPO),啄木鸟取食时停留在树的部位,划分为主干(记为1)、侧枝(记为2);
(16) 取食方式(DRT),划分为凿干(记为1)、扒树皮(记为2);
(17) 单树取食时间(FDT),啄木鸟取食时在同一棵树上消耗的时间,自观察到啄木鸟取食或听见凿击声时开始计时,精确到分钟。
对照样方中,使用同样方法收集和记录上述(4) —(9)共6项生境因子数据。
1.3 数据分析方法
先采用多变量回归树(Multivariate regression trees, MRT)分析3种啄木鸟取食生态位是否存在差异,再用多分类逻辑斯谛回归(multinomial Logistic regression, MLR)判断3种啄木鸟在哪些取食生境因子利用和取食行为特征上存在差异。对于存在差异的因子,使用两因子方差分析(two-ways ANOVA)或Bailey′s方法,进行进一步的分析,具体方法如下。
1.3.13种啄木鸟取食生境及取食行为特征多变量回归树
将鸟种和性别变量数据作为响应变量矩阵,余下的变量数据除去鸟种、性别和单树取食时间后,使用R语言环境下的vegan包[24]的decostand函数转换为标准化矩阵,作为自变量矩阵。将这两个矩阵使用R语言环境下的MVPARTwrap包[25]的mvpart函数在交互环境下计算并生成多变量回归树[26]。
1.3.23种啄木鸟取食生境和行为特征的多分类逻辑斯谛回归分析
以鸟种为因变量,以除性别、取食时刻、单树取食时间以外的其他变量为自变量,使用R语言环境下的nnet包[27]的multinom函数进行多分类逻辑斯谛回归[26],并采用逐步回归的方法进行模型优化。自变量中的分类变量全部转换为因子型(factor)。对进入最优模型的变量,使用似然比检验(likelihood ratio test)进行显著性检验[26]。检验显著的变量进入后续分析。
1.3.33种啄木鸟取食行为节律分析
使用R语言环境下的ggridges包[28]的geom_density_ridge函数,依据收集的取食时刻数据,分别计算3种啄木鸟在不同时刻取食的概率密度函数,制作叠嶂图(ridgeline plot)展示3种啄木鸟之间的区别。
1.3.43种啄木鸟对取食生境因子的偏好差异或取食行为特征的表现差异
对于郁闭度、树密度、死树密度、取食树高、取食树胸径、单树取食时长等定量数据,使用两因子方差分析(two-ways ANOVA),以鸟种和性别为自变量,以相应变量为因变量进行统计检验。
对于因子水平数超过3个的取食生境和取食行为特征变量,包括林型、坡位、优势树种、取食树种和取食部位,以对照样方为参照,使用Bailey′s方法分析3种啄木鸟对相应变量的选择或行为表现偏好[23]。
对于取食树状态、取食部位和取食方式3个两水平取食行为特征变量,如有需要,采用卡方检验(χ2test)比较不同啄木鸟之间的差别。
全部数据分析在R3.4.4环境下,使用R语言完成[29]。除特别说明外,数据以平均数±标准误(mean±SE)表示,统计检验显著性水平设定为α=0.05。
2 结果
野外调查共收集啄木鸟取食数据312组,其中黑啄木鸟73组,三趾啄木鸟97组,大斑啄木鸟142组。
2.1 3种啄木鸟取食生态位的分离
交叉验证提示,4分支是多变量回归树(MRT)的最优分支数。MRT结果显示,3种啄木鸟分别聚成三支,提示基于参与分析的取食生境和取食行为特征变量,3种啄木鸟的取食生态位有明显差异(图1)。
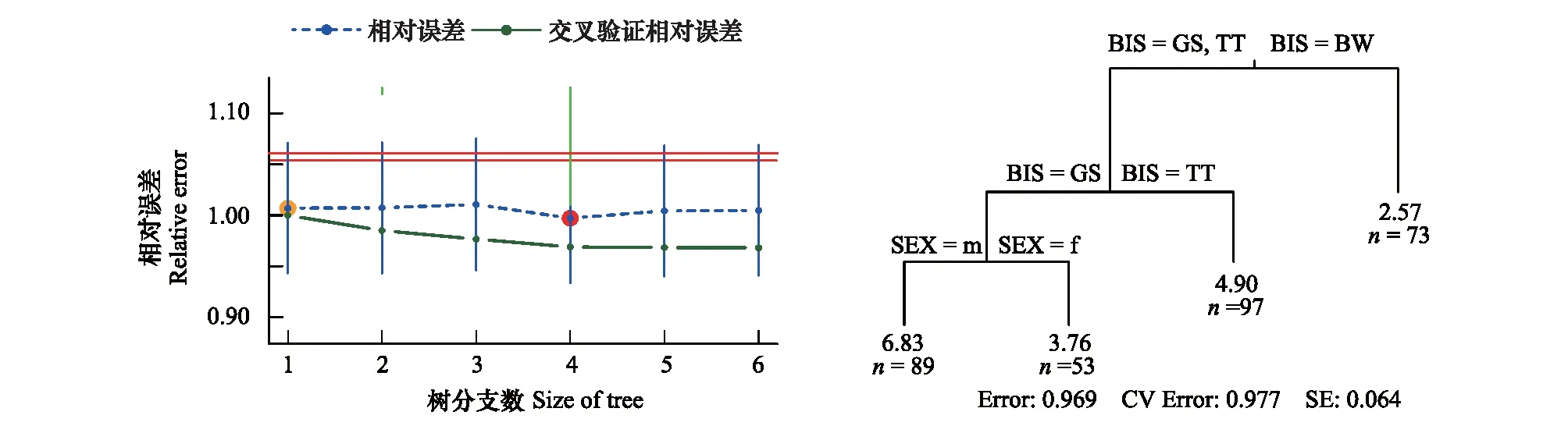
图1 3种啄木鸟取食生境及行为特征多变量回归树分析结果Fig.1 Multivariate regression tree on forage habitat and behavior characters of three woodpecker species红点提示最优分支数,多变量回归树BIS:鸟种bird species BW:黑啄木鸟black woodpeckers (Dryocopus martius); TT:三趾啄木鸟three-toed woodpeckers (Picoides tridactylus); GS:大斑啄木鸟great spotted woodpeckers (Dendrocopos major); m:雄性male; f:雌性female
与此同时,逐步多分类逻辑斯谛回归结果提示,全部参与分析的取食生境和取食行为特征变量,都对3种啄木鸟取食生态位的分离有显著影响(表1)。
2.2 3种啄木鸟的取食行为节律差异
取食时刻的概率密度函数提示,3种啄木鸟的取食行为节律有明显差异(图2)。黑啄木鸟表现为双峰型,分别在清晨和黄昏出现一个取食活动高峰。大斑啄木鸟和三趾啄木鸟则全天活动,13:00—14:00分别达到取食活动最高峰。

表1 3种啄木鸟取食生境因子逐步多分类逻辑斯谛回归结果
FOT: 林型forest type; SLP: 坡位 slope position; DTS: 优势树种 dominant tree species; CRD:郁闭度 crown density; TRD: 树密度 tree density; SND: 死树密度 snag density; FTS: 取食树种 forage tree species; HET: 树高 height of the forage tree; DBH: 胸径 diameter at breast height of forage tree;TSI: 树状态 forage tree situation; FHE: 取食高度 forage height;FPO: 取食位置 forage position; DRT:凿取方式 drill type
2.3 3种啄木鸟对取食生境特征的选择差异
Bailey′s分析结果显示,3种啄木鸟对林型、坡位、取食地优势树种、取食树种和取食部位的偏好有显著的差异(表2)。大斑啄木鸟取食范围比较广,除明显偏好在阔叶树上取食、偏好在冠层活动而回避在树基部活动外,对其他生境因子的不同水平表现为随机选择。三趾啄木鸟和黑啄木鸟的取食生境偏好比较一致,但三趾啄木鸟取食范围更为广泛,体现在对林型的选择上,除偏好原始林外,对其他林型都表现为随机选择,而黑啄木鸟从未在人工林中被发现(表2)。
3种啄木鸟的取食树高,无论在不同鸟种间还是同种鸟的不同性别间都没发现显著差异(Two-way ANOVA, 鸟种间,F2,308=1.466,P=0.232; 两性间:F1,308=0.229,P=0.632)。而取食生境的郁闭度、树密度、死树密度及取食树胸径间则有显著差异(图3)。黑啄木鸟和三趾啄木鸟多活动在郁闭度相对较高的生境中(BW, 0.67±0.02; TT, 0.62±0.02; GS, 0.53±0.02),大斑啄木鸟则多活动在树密度相对较高的生境中(BW, 39.9±1.2; TT, 37.4±1.3; GS, 43.3±1.7),三趾啄木鸟活动的生境中死树密度最低(BW, 3.8±0.1; TT, 3.0±0.1; GS, 3.6±0.2),而大斑啄木鸟选择的最食树胸径相对最大(BW, 35.8±0.7; TT, 39.8±1.0; GS, 40.7±0.9)。

表2 3种啄木鸟对取食生境特征的选择

图2 3种啄木鸟取食行为节律 Fig 2 Forage behavior rhythms of three woodpecker species蓝色部分: 12:00前的取食活动概率blue fill: forage behavior probability before 12:00; 红色部分: 12:00后的取食活动概率 red fill: forage behavior probability after 12:00; 两种颜色交叠部分: 12:00前后的活动概率(红色值和蓝色值的和) overlap of blue and red fill: forage behavior probability around 12:00(sum of the value indicated by red and blue plot)
BW:black woodpeckers (Dryocopusmartius); TT:three-toed woodpeckers (Picoidestridactylus); GS:great spotted woodpeckers (Dendrocoposmajor); Ex.Rat:excepted ratio; O.N:observation numbers; LL:lower limit; UL:upper limit; SL:selectivity; +:偏好选择 (observed usage is significantly higher than expected); -:避免选择 (observed usage is significantly lower than expected);o:随机利用 (observed usage in proportion to its availability)
2.4 3种啄木鸟取食行为的差异
黑啄木鸟多在死树上取食,大斑啄木鸟则多在活树上取食,而三趾啄木鸟的取食树则活、死树各半(图4)。与黑啄木鸟和三趾啄木鸟多在树干上取食不同,大斑啄木鸟有43.7%的比例在树枝上凿取食物(图4)。三趾啄木鸟多数时间都是通过扒去树皮获取食物,而黑啄木鸟和大斑啄木鸟则绝大多数情况下通过在树干上凿洞获取食物(图4)。
2.5 3种啄木鸟单树取食时长的差异
3种啄木鸟的单树取食时长存在显著差异,黑啄木鸟和三趾啄木鸟经常在同一株上取食超过20 min,而大斑啄木鸟则经常变换取食树(图5)。
3 讨论
3.1 研究方法的使用
传统的研究,多采用计算生态位宽度和生态位重叠指数的方法,分析物种间的生态位分离程度[15-16],取得了很好的结果。但是,这类指数在计算过程中,实际上将各个变量进行了平均化处理,大量丢失了变量所包含的信息。采用近年来发展出的多变量回归树(MRT)、多分类逻辑斯谛回归(MLR)等多变量数据分析技术(multivariate data analysis methods)[26],最大限度地利用了数据中所包含的信息,提高了识别生态位分异的敏感性。MRT和MLR的结果一致显示,在所调查的取食生境和取食行为特征上,3种啄木鸟的取食生态位出现了明显的分离(图1,表1)。

图3 3种啄木鸟的取食生境数量特征差异Fig.3 Differences of forage habitat quantitative characters among three woodpecker species方差分析结果 two-ways ANOVA Results: 胸径, 物种间, F2,308=13.056, P=3.609×10-6, 两性间, F1,308=0.552, P=0.458; 树密度, 物种间, F2,308=3.781, P=0.024, 两性间, F1,308=1.196, P=0.275; 死树密度, 物种间, F2,308=3.588, P=0.029, 两性间, F1,308=0.418, P=0.519; 胸径, 物种间, F2,308=6.738, P=0.001, 两性间, F1,308=0.447, P=0.504
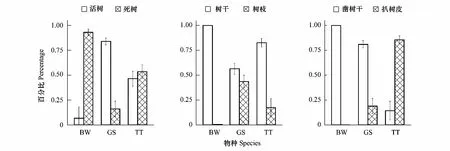
图4 3种啄木鸟的取食行为差异Fig.4 Differences of forage behavior characters among three woodpecker species

图5 3种啄木鸟的单株树上取食消耗时间 Fig.5 Differences of forage duration per tree among three woodpecker species方差分析结果two-ways ANOVA Results: 物种间, F2,308=190.274, P<2×10-16; 两性间, F1,308=0.004, P=0.950
需要指出的是,多变量数据分析技术与传统的统计检验方法在方法论上有很大的差别,两类方法有时会出现表面上相反的结果。比如本研究中,表1显示取食树高在区别3种啄木鸟有着显著的作用,但two-way ANOVA分析的结果显示,3种啄木鸟的取食树高没有显著差别。出现这种矛盾的原因在于,黑啄木鸟和三趾啄木鸟多在原始云、冷杉林中活动(表2),且多在云、冷杉树上活动,这些树木是取食生境中的最高树。而大斑啄木鸟虽然可以活动在不同生境中,但选择在胸径较大的树上取食(图2,图3),而这些树同样是取食生境的最高树,3种啄木鸟取食树也因此没有显著的高度差别。但是,这些树出现在不同的坡位、不同的林型中,这一点被MLR方法识别了出来,用于区别不同的啄木鸟。同样地,进一步的数据分析也没有发现图1中所体现的两性间的差异,3种啄木鸟两性间在取食生境和取食行为上没有显著差异(图2—图5)。
3.2 取食活动节律的差异
3种啄木鸟的体形差别很大,黑啄木鸟最大,体重可达350 g,而三趾啄木鸟和大斑啄木鸟的体重通常不足黑啄木鸟的四分之一。体重的差别,导致后两种啄木鸟冬季比黑啄木鸟要消耗相对更多的能量。正因为如此,黑啄木鸟多在日出后和黄昏前取食,中午则很少取食和活动(图2),而三趾啄木鸟和大斑啄木鸟则全天取食(图2)。这与胡加付等关于大斑啄木鸟的研究是一致的[30],但三趾啄木鸟和黑啄木鸟尚无相关的研究报道。
3.3 3种啄木鸟取食生态位的差异
与高玮等的研究结论相一致[15-16],同域分布的啄木鸟对取食生境的选择和取食行为特征上,存在着明显的差异(表1,图1)。
黑啄木鸟和三趾啄木鸟的取食生境很相近,都偏好活动于原始林中(表2),偏好于在云、冷杉林中取食(表2)。这些林地多位于谷地(表2),这样的生境中死树的密度也相对较高(图3)。由于优势树种是云、冷杉,这两种啄木鸟多在云、冷杉树的树干上取食(表2)。二者不同的是,除了活动时间相异外,黑啄木鸟的喙粗大强直,多通过在树干或倒木上凿挖深坑,控取树干深处的昆虫幼虫,而三趾啄木鸟喙幼弱,只能扒取树皮下的昆虫(图4)。与此相适应,很可能是因为死树的树干较软,更容易凿挖[31],且含有更多食物,黑啄木鸟多在死树上取食,而三趾啄木鸟取食树则死活参半 (图4)。也正因为需要开挖很大的凿洞,黑啄木鸟在同一棵树上花费的时间很长(图5)。附加的调查显示,同一只黑啄木鸟会连续几天在同一棵树上取食,直至将那棵树凿得千疮百孔(另文报道)。此外,三趾啄木鸟利用的生境也较黑啄木鸟稍广泛(图3,图4),这很可能跟三趾啄木鸟仅能扒取树皮下的昆虫,食物资源更为紧张有关,更广泛的取食生境能够提供更多的食物。
与已有的研究相一致,大斑啄木鸟是生境利用泛化者[32],能够在不同的环境中取食(表2,图3,图4),因此可以获得更为广泛的食物来源[7,33],这很可能是大斑啄木鸟遇见率最高的原因[2,34-35]。3种啄木鸟中,大斑啄木鸟的取食生境郁闭度最低、树密度最高,而取食树胸径最大(图3),这个结果看似矛盾实则不然。出现这样的结果,正是因为大斑啄木鸟可以在次生林、人工林中活动,这些林地恰恰树木密度大但郁闭度低,处于演替早期,死树密度低(图3),大斑啄木鸟可以选择在生境中更大的树上(图3),而且是活树上完成取食活动(图4),获得更多的食物,提高取食效率。
调查中经常能看到黑啄木鸟和三趾啄木鸟出现在同一样方,但由于取食部位和取食方式的不同,从未见到二者互相驱离的现象。3种啄木鸟取食生态位的分异,无异减轻了彼此之间的竞争,使它们能够更有效地利用有限的食物资源,从而实现同类物种的同域共存[2, 7]。
纵观有关啄木鸟同域共存的研究,大斑啄木鸟是最常见的研究对象[2, 7,33-35],这与它广泛的分布范围有关,三趾啄木鸟的研究很少[18],黑啄木鸟几乎没有。不仅有关啄木鸟物种共存的研究不多,有关啄木鸟的研究整体都很匮乏。根据检索,目前有关啄木鸟的文献不超过700篇,这不仅与254种鸟种的数量不相称,更与啄木鸟在森林生态系统中的地位不相称,啄木鸟的生态学研究有待深入。
猜你喜欢
幼儿100(2023年18期)2023-05-29 08:34:36
区域治理(2023年6期)2023-03-02 14:34:54
中国农学通报(2022年32期)2022-11-28 05:01:16
湖南大学学报(自然科学版)(2022年8期)2022-09-02 04:22:42
课堂内外·小学版(低年级)(2022年4期)2022-06-24 09:47:10
大众摄影(2021年3期)2021-03-22 02:45:43
——以北长山岛为例
海洋开发与管理(2020年9期)2020-10-13 13:31:22
现代农业科技(2020年10期)2020-06-04 09:13:31
孩子(2019年8期)2019-08-26 05:43:32
十几岁(2016年1期)2016-04-09 07:20:11
