
不同模式的低氧训练对骨骼肌肌球蛋白合成及其mTOR信号通路的影响
2019-01-18 03:22:04杨加玲王海军
中国运动医学杂志 2018年12期
杨加玲 王海军
江苏省连云港师范高等专科学校(连云港 222006)
高住高练(living high-training high,HiHi)和高住低练(living high-training low,HiLo)是低氧训练的两种常见模式[1],HiHi是在低氧环境中生活和训练,而Hi-Lo是在低氧条件下生活,但在常氧环境下进行运动训练。研究表明,在高原低氧和运动缺氧的双重刺激下,机体蛋白质的降解率会大于其合成率,导致机体蛋白质丢失,引起肌肉萎缩,肌力下降[2,3],从而影响了高原训练的效果。与高原训练相比,HiLo可使骨骼肌蛋白质丢失的程度减少,在一定程度上可维持骨骼肌的收缩力量[1]。但是,在模拟高原训练或HiLo过程中,骨骼肌蛋白质含量降低的机制目前还不完全清楚。有研究表明,在骨骼肌蛋白质合成的调节过程中,PI3K/Akt/mTOR(雷帕霉素靶蛋白)信号通路具有非常重要的促进作用[4,5],抑制PI3K/Akt/mTOR信号通路的活性可明显促进骨骼肌的废用性萎缩,而PI3K/Akt/mTOR信号通路的激活可使骨骼肌废用性萎缩的程度明显下降[4]。但是,PI3K/Akt/mTOR及其下游信号通路在调节低氧运动训练过程中骨骼肌合成中的作用目前研究很少。本研究对大鼠进行低氧暴露和/或大强度的运动训练,6周后测定大鼠腓肠肌中总蛋白(Pro)和肌球蛋白(Myo)含量,同时测定腓肠肌中PI3K、Akt含量以及mTOR、真核细胞延伸因子2(eukaryotic elongation factor 2,eEF2)和真核细胞翻译起始因子4E结合蛋白1(eukaryotic translation initiation factor 4E binding protein 1,eIF4E-binding proteins,4EBP-1)的mRNA表达,探讨不同模式低氧训练过程中骨骼肌蛋白质合成代谢的变化及其调节机制,旨在促进低氧训练科学化。
1 对象与方法
1.1 实验对象和分组
体重130~160 g的雄性SD大鼠50只,由江苏大学实验动物中心提供,许可证号为SCXK(苏)2013-0011。在室温20~22℃的环境下分笼饲养,每笼5只,喂饲国家标准啮齿类动物固体饲料,自由饮水。大鼠在3天的适应性饲养后,随机被分成常氧对照组(NC组,n=10)、低氧暴露组(HC组,n=10)、常氧训练组(NE组,n=10)、模拟高原训练组(HE组,n=10)和高住低练组(HiLo组,n=10)5组。
1.2 低氧与训练方案
NC和HC组分别在常氧和低氧环境中生活,不进行运动训练;NE和HE组分别在常氧和低氧环境中生活和训练;HiLo组在低氧环境中生活12 h,其余时间在常氧下生活和运动训练。实验所需要的低氧环境由The MAG-10 Mountain Air Generator模拟,在第1周内将低氧舱中氧浓度从模拟海拔高度1600 m逐渐增加到3500 m,最终氧浓度为13.6%。NE、HE和HiLo组在下午进行负重游泳训练,每周6天,训练6周。在第1周内将常氧下和低氧下训练大鼠的负重逐渐增加到体重的1.9%(相当于大鼠90%乳酸阈强度)[6]和1.7%[7],游泳时间渐增到90 min,以后按照此运动强度和时间进行训练。游泳池为内壁光滑的长方体塑料水桶(120 cm×80 cm×70 cm),游泳池水的深度保持身高的2倍以上,温度维持在30~33℃之间。
1.3 取材
末次运动训练后第2天上午进行实验取材,取材前禁食禁水12 h,腹腔注射20%的乌拉坦溶液对大鼠进行麻醉后,快速分离腓肠肌,将筋膜和脂肪去除,装于冻存管放入液氮中速冻,最后保存至-80℃冰箱。
1.4 指标测试
腓肠肌Myo、PI3K、Akt含量采用ELISA测定,总蛋白含量采用BCA法测定,试剂盒由南京森贝伽生物科技有限公司提供,仪器为multiskan全波段酶标仪。4EBP-1、eEF2和mTOR mRNA采用Real-time PCR法[7],RNAiso Plus和cDNA合成试剂盒购自TaKaRa公司,SYBR Green Master(ROX)试剂盒购自Roche Diagnostics,测定仪器为ABI 7500 RT-PCR仪。由上海生物工程有限公司合成引物,序列见表1。

表1 各种指标和GAPDH的引物序列
1.5 数据分析
使用SPSS 22.0统计软件进行处理,实验结果用x±s表示,NC、HC、NE和HE 4组之间进行双因素方差分析,HiLo、NC、HE 3组之间进行单因素方差分析和多重比较,P<0.05有显著性差异,P<0.01有极显著性差异,。
2 结果
2.1 各组大鼠腓肠肌总Pro含量、Myo含量的变化
由表2、表3可知,无论是低氧暴露还是大强度的运动训练,均使腓肠肌Pro含量显著降低(P<0.05,P=0.01),Myo含量也显著下降(P<0.01,P=0.05),低氧暴露与运动训练虽然对降低腓肠肌Pro含量无显著性交互作用,但对降低Myo含量有显著性的交互作用(P<0.05);HiLo组腓肠肌Pro和Myo含量显著低于NC组,但显著高于HE组(P<0.05)。
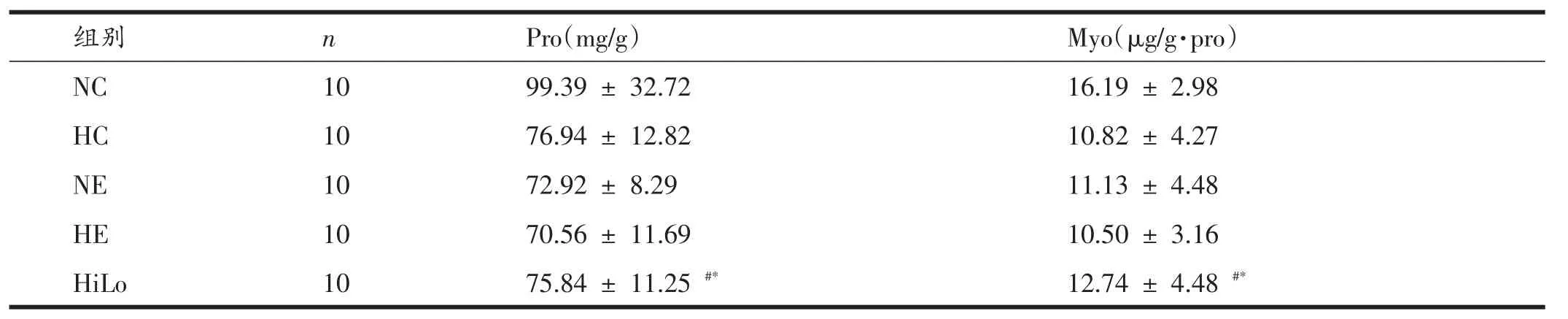
表2 各组腓肠肌Pro、Myo含量

表3 各组腓肠肌Pro、Myo含量的双因素方差分析
2.2 各组实验大鼠腓肠肌4E-BP1 和eEF2 mRNA表达量的变化
由表4、表5可知,对NC、NE、HC和HE组4组之间进行双因素方差分析,低氧暴露使腓肠肌4E-BP1 mRNA表达量显著升高(P<0.05),eEF2mRNA表达有所降低,但无显著性差异;而运动训练使腓肠肌4E-BP1 mRNA表达量显著升高,eEF2mRNA表达显著降低(P<0.05);低氧和大强度运动训练对4EBP-1 mRNA表达的进一步升高无显著性交互作用,而对降低腓肠肌eEF2 mRNA表达有显著性交互作用(P<0.05);HiLo组腓肠肌4E-BP1mRNA的表达显著高于NC组,虽低于HE组,但无显著性差异;eEF2mRNA表达与NC组相比无显著性升高,但是显著高于HE组(P<0.01)。

表4 各组腓肠肌4E-BP1和eEF2的mRNA表达

表5 各组腓肠肌4E-BP1和eEF2 mRNA表达的双因素方差分析
2.3 各组大鼠腓肠肌PI3K、Akt含量和mTORmRNA表达的变化
由表6、表7可知,对NC、NE、HC和HE组4组之间进行双因素方差分析发现,低氧暴露使腓肠肌PI3K、Akt含量和mTOR mRNA表达显著降低(P<0.01,P<0.05,P<0.01),运动训练使PI3K含量和mTOR mRNA表达显著降低(P<0.05,P<0.01),低氧暴露与运动训练对降低腓肠肌PI3K和Akt含量无显著性交互作用,但对降低mTOR mRNA表达具有显著性的交互作用(P<0.01);HiLo组腓肠肌PI3K、Akt含量和mTOR mRNA表达与NC组相比无显著性变化,但PI3K含量和mTOR mRNA表达显著高于HE组(P<0.05)。
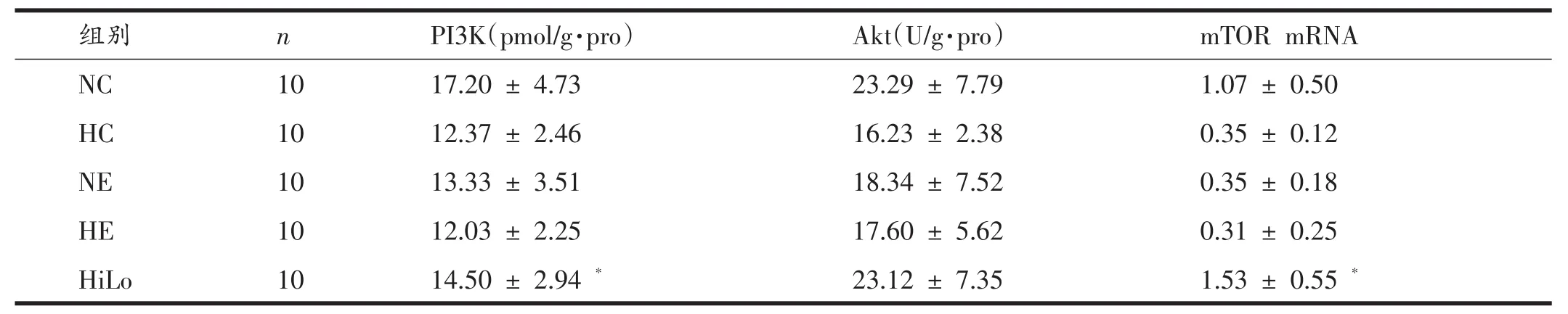
表6 各组腓肠肌PI3K、Akt的含量和mTOR的mRNA表达

表7 各组腓肠肌PI3K、Akt含量和mTOR mRNA表达的双因素方差分析
3 讨论
在组成骨骼肌的成分中,蛋白质是最重要的,其中主要的结构蛋白和收缩蛋白是肌球蛋白,其含量的降低不仅可降低骨骼肌的收缩速度,还可减小骨骼肌的收缩力量。为了进一步探讨高原训练过程中导致骨骼肌蛋白质下降的主要原因,本研究通过交互设计的实验方案对大鼠进行低氧暴露和/或运动训练,6周后检测了腓肠肌Pro、Myo含量。结果发现,无论是6周的大负荷游泳训练还是低氧暴露均显著降低了大鼠腓肠肌Pro和Myo含量,虽然低氧暴露和运动训练两个因素对腓肠肌Pro含量的减少无显著性交互作用,但对降低Myo含量具有显著性的交互作用。从而说明,机体长期暴露在低氧环境中或进行大负荷运动训练时,骨骼肌蛋白质的丢失均增加,而在低氧环境中进行大负荷运动训练时骨骼肌收缩蛋白的丢失更加明显。然而HiLo组大鼠腓肠肌Pro和Myo含量虽然显著低于NC组,但均显著性高于HE组。因此,与传统的高原训练相比,HiLo可以降低大鼠骨骼肌蛋白质丢失的程度。
有研究证实,PI3K/Akt/mTOR信号通路是以mTOR为中心的信号转导途径,在细胞生长过程中发挥着关键作用[8,9]。它通过调节核糖体S6激酶(p70S6K)、翻译抑制蛋白4E-BP1、翻译起始因子eIF4E和eIF4G及翻译延长因子eEF2等下游信号因子调节蛋白质的生物合成。其中,eEF2属于翻译延伸过程中的一种重要的细胞因子,它能与核糖体结合,促进mRNA翻译延长过程。eIF4E与eIF4A和eIF4G结合形成翻译起始所必须的eIF4F复合物。由于4E-BP1能和eIF4G竞争性地与eIF4E结合,可阻碍eIF4F复合物的形成,故是eIF4E的抑制因子。因此,4E-BP1是eIF4F复合物形成过程中最为关键的调节因子,是mTOR直接作用的底物之一[10,11]。PI3K/AKT/mTOR信号通路的活化,可能会下调4EBP-1和上调eEF2的表达,进而促进蛋白质的合成[12,13]。
有研究表明,低氧可以通过抑制骨骼肌IGF-1的分泌或阻断IGF-1介导的Akt/mTOR的激活[14-16],从而抑制mTOR调节下游的4EBP-1和eEF2(真核细胞延伸因子2)来降低蛋白质合成[17],而eEF2的磷酸化与去磷酸化的平衡最终影响其活性[18]。运动时AMPK激活,从而激活eEF2激酶的活性,引起eEF2磷酸化,使eEF2活性降低,可抑制蛋白质的合成[18,19];运动后mTOR激活,能使eEF2去磷酸化,使eEF2活性升高,促进蛋白质的合成。目前不少研究证实,力量训练可以激活骨骼肌Akt/mTOR信号通路促进蛋白质的生物合成,从而引起骨骼肌体积的增大[20-22]。而耐力性运动训练对骨骼肌Akt/mTOR信号通路的影响目前研究结果并不一致,且可能与运动强度有关。低强度的有氧耐力性运动训练并不能使骨骼肌中mTOR活性发生变化[23],而进行不习惯或大强度的耐力性运动可以通过提高AMPK活性抑制骨骼肌中mTOR的活性[20]。但是有研究发现,28天的耐力性运动训练可激活大鼠骨骼肌Akt/mTOR/p70S6K信号通路,促进骨骼肌蛋白质的生物合成[24,25]。然而,低氧运动对Akt/mTOR及其下游信号通路影响的报道甚少。有研究证实,HiLo对骨骼肌Akt/mTOR信号通路具有一定的抑制作用,在一定程度上可以抑制蛋白的生物合成[24]。但也有研究发现,低氧暴露可通过抑制骨骼肌mTOR/p70S6K信号通路抑制肌肉蛋白质的合成,在低氧中进行适量的耐力运动锻炼在一定程度上可削弱低氧对mTOR/p70S6K信号通路的抑制作用[25]。有研究发现,2个月的运动训练虽然能够增加血氧正常和血氧低下的慢性阻塞性肺疾病(COPD)患者的运动能力,但运动训练只能增加血氧正常COPD患者骨骼肌中柠檬酸合成酶和乳酸脱氢酶活性、肌纤维横截面积和毛细血管纤维比值。与此同时,血氧正常COPD患者骨骼肌中Akt(Ser473)、GSK-3β(Ser9)和p70S6k(Thr389)的磷酸化在运动训练后无显著性增加,但在血氧低下的COPD患者中显著降低[16],从而进一步说明运动训练在低氧和常氧下对骨骼肌蛋白质生物合成的影响不同。本研究发现,6周的低氧暴露在显著降低大鼠腓肠肌PI3K、Akt含量和mTOR mRNA表达的同时,显著升高4E-BP1 mRNA表达量,而eEF2 mRNA表达无显著性降低;运动训练能显著降低PI3K含量以及mTOR和eEF2 mRNA表达,显著升高4EBP1 mRNA表达量;且低氧暴露与运动训练对降低mTOR和eEF2 mRNA表达具有显著性的交互作用;与NC组相比,HiLo组除了腓肠肌4E-BP1 mRNA的表达显著升高外,PI3K、Akt含量和mTOR mRNA表达均无显著性变化,但HiLo组PI3K含量、mTOR和eEF2的mRNA表达显著高于HE组。
长时间的低氧暴露和大强度的运动训练,由于骨骼肌能量的大量消耗,一方面激活AMPK,另一方面促进皮质酮的分泌[26],抑制睾酮和IGF-1的分泌[2],从而抑制PI3K/Akt/mTOR信号通路,显著上调4EBP-1和下调eEF2 mRNA表达。而低氧联合大强度运动训练在一定程度上对骨骼肌PI3K/Akt/mTOR及其下游信号通路的抑制作用比单纯的低氧暴露或大强度运动训练更大。但是由于HiLo中,低氧暴露的时间比模拟高原训练要短,从而使HiLo对骨骼肌PI3K/Akt/mTOR及其下游信号通路的抑制作用比模拟高原训练要小。因此,在低氧训练中如何采取低氧暴露时间和运动强度的合理组合,在最大程度上减轻低氧暴露引起的蛋白质丢失,对于提高运动员的运动能力有着非常重要的意义。
4 结论
4.1 模拟高原训练中,低氧和大强度的运动训练均能通过抑制骨骼肌PI3K/Akt/mTOR及其下游信号通路,降低骨骼肌蛋白质含量。低氧联合大强度运动训练对骨骼肌PI3K/Akt/mTOR及其下游信号通路以及蛋白质合成的抑制作用在一定程度上比单纯的低氧暴露或大强度运动训练更大。
4.2 高住低练能够减轻骨骼肌中PI3K/Akt/mTOR及其下游信号通路的抑制程度,减轻抑制骨骼肌蛋白质合成的程度,对于防止低氧引起的骨骼肌蛋白质丢失具有重要作用。
猜你喜欢
分子诊断与治疗杂志(2023年8期)2023-09-07 12:00:34
天津医科大学学报(2021年3期)2021-07-21 09:03:42
中成药(2018年10期)2018-10-26 03:41:40
国外医药(抗生素分册)(2016年5期)2016-07-12 14:25:34
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
中国运动医学杂志(2016年3期)2016-07-10 12:07:23
磁共振成像(2015年5期)2015-12-23 08:52:53
医学研究杂志(2015年5期)2015-06-10 06:43:26
实验动物与比较医学(2014年5期)2014-02-28 14:53:12
中华骨与关节外科杂志(2014年4期)2014-01-22 12:14:24
