
饲料亮氨酸水平对幼草鱼生长、饲料利用及体成分的影响
2018-12-29 07:27:28黄爱霞孙丽慧陈建明沈斌乾姜建湖
饲料工业 2018年2期
■黄爱霞 孙丽慧 陈建明沈斌乾 姜建湖
(浙江省淡水水产研究所农业部淡水渔业健康养殖实验室浙江省鱼类健康与营养重点实验室,浙江湖州313001)
蛋白质是水生动物饲料中成本最高的部分,鱼类对蛋白质的需求,实际上就是对氨基酸的需求。氨基酸不仅是蛋白质的构成物质,还在机体生理代谢过程中发挥着重要的作用[1-2]。因此鱼类氨基酸需要量的确定,对配制营养均衡、廉价高效、低污染的配合饲料有非常重要的指导意义[3]。亮氨酸是鱼类的必需氨基酸,研究表明,在饲料中添加适量的亮氨酸,可以促进摄食和提高饲料效率,进而提高鱼类的生长性能。
草鱼(Ctenopharyngodon idellus Valenciennes)是我国著名的淡水“四大家鱼”之一,早期对其营养需求的研究主要集中在蛋白质、脂肪和碳水化合物等方面[4-7],其研究成果对草鱼配合饲料的开发和应用起到了极大的推进作用。近些年对其必需氨基酸需求的研究已有报道,但并不完善,目前仅见对精氨酸[8]、赖氨酸[9]、苏氨酸[10]、异亮氨酸[11]、苯丙氨酸[12]、缬氨酸[13]和亮氨酸[14]等需求量的报道。其中Deng等[14]研究了饲料亮氨酸水平对草鱼大规格鱼种[(295.85±2.07)g]的影响,而本试验则是通过调节饲料中亮氨酸水平探讨亮氨酸对草鱼幼鱼(2.25 g)生长、饲料利用等指标的影响,从而得出其适宜的亮氨酸需求量,为开发氨基酸平衡的草鱼幼鱼高效配合饲料提供科学依据。
1 材料与方法
1.1 试验鱼
试验鱼为浙江省淡水水产研究所试验基地同塘培育的当年夏花鱼种。选取规格整齐的健壮个体,放入已经编号的水族箱中,用试验基础饲料驯养一周,待试验鱼摄食较好后,正式开始试验。将1 050尾试验鱼随机分成7组,每组3个重复,每个重复放50尾鱼。试验鱼初始体重为2.25 g,各组之间无显著差异(P>0.05)。
1.2 试验饲料
试验饲料以鱼粉和晶体氨基酸混合物为蛋白源;氨基酸模式参照测定的试验草鱼全鱼40%粗蛋白质中氨基酸含量(亮氨酸除外);用晶体亮氨酸调节饲料亮氨酸水平;用天冬氨酸和谷氨酸(1∶1)调整百分比。试验饲料亮氨酸实测值为9.65、12.05、14.98、17.31、20.06、22.88、25.60 g/kg(占饲料的质量分数),试验饲料设计配方的原料组成及营养水平见表1。试验饲料制作时,先将原料粉碎,并过60目筛,再按比例混合并搅拌均匀后,用绞肉机挤压成型,制成直径1 mm左右的硬颗粒。于50℃烘干,冷却后入塑料袋包装,置于4℃冰箱冷藏备用。试验饲料必需氨基酸组成见表2。

表1 饲料配方组成及营养水平(g/kg干饲料)
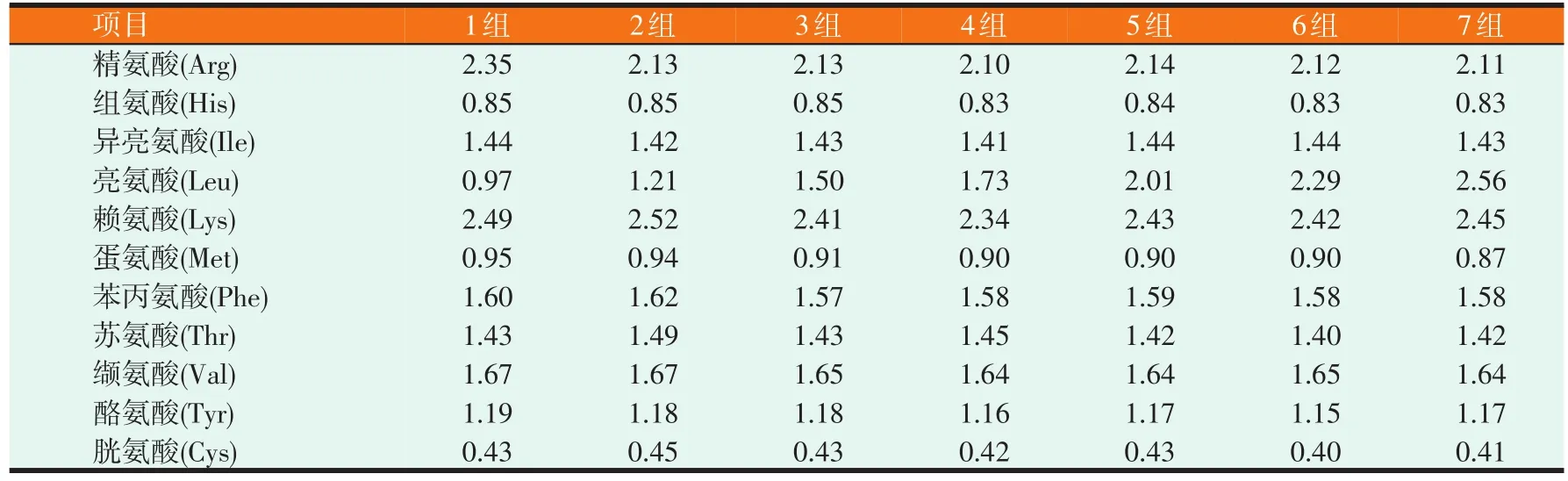
表2 饲料必需氨基酸组成分析(干物质基础,%)
1.3 饲养条件
试验于2015年8月5日至10月14日在浙江省淡水水产研究所试验基地进行。养殖系统为室内玻璃缸循环水养殖系统,养殖水体(0.8 m×0.5 m×0.4 m),缸内配有生物过滤装置和气石。试验用水为充分曝气的自来水,日换水量为总水体的一半,连续充气。采用饱食投喂法,投饲时间为上午8:00、中午12:00和下午4:00。试验期间,水温25~30 ℃;水质:pH值7.4~8.0,溶氧>5.0 mg/l,氨氮0.14~0.20 mg/l,亚硝酸盐0.02~0.05 mg/l。试验期间每天记录投饲量。
1.4 取样及分析
试验开始时随机取20尾鱼作为初始鱼样。饲养试验结束后,试验鱼停饲24 h称终末体重。随机取鱼10尾为一混合样,用于测定试验鱼的全鱼组成及氨基酸组成;另取5尾鱼称量全长和体重,于冰盘上解剖取内脏和肝脏称重,用于计算肥满度、脏体指数和肝体指数,肝脏先放入液氮罐中速冻,再保存到-20℃冰箱中用于分析肝脏谷丙转氨酶(ALT)和谷草转氨酶(AST)活性。
水分、粗蛋白质、粗脂肪和灰分的测定参照AOAC的方法[15]:水分测定采用失重法;粗蛋白质测定采用凯氏定氮法;粗脂肪测定采用索氏抽提法;灰分测定采用马福炉550℃焚烧法;样品氨基酸组成测定采用酸水解法,通过6 mol/l的浓盐酸110℃真空水解24 h的预处理后,用氨基酸分析仪测定;肝脏ALT、AST活性的测定按照南京建成生物工程研究所提供的测试盒说明书采用赖氏法进行测定。
鱼体增重(g)=末体重-初体重
饲料效率=(末体重-初体重)/饲料摄入量
成活率(%)=100×(末尾数/初尾数)
蛋白质积累率(%)=100×(末体重×末草鱼蛋白质含量-初体重×初草鱼蛋白质含量)/(饲料摄入量×饲料蛋白质含量)
脏体比(%)=100×(肝脏重/鱼体重)
肝体比(%)=100×(内脏重/鱼体重)
肥满度(g/cm3)=鱼体重(g)/[鱼体长(cm)]3
1.5 数据处理及统计分析
试验数据采用“平均数±总标准误”表示,采用SPSS16.0 for Windows分析软件(One-way ANOVA)对所得试验据进行单因素方差分析,若有显著差异再做SNK多重比较确定组间差异性。显著水平P采用0.05,若P<0.05为差异显著。采用折线回归分析模型来拟合鱼体增重与亮氨酸水平之间的关系,求得草鱼幼鱼亮氨酸的最佳需求量。
2 结果与分析
2.1 草鱼幼鱼生长情况(见表3)
由表3可知,饲料亮氨酸水平对草鱼幼鱼体增重、饲料效率和蛋白质积累率均有显著影响(P<0.05)。体增重随饲料亮氨酸水平的升高而增加,当饲料亮氨酸水平达14.98 g/kg时,继续增加饲料亮氨酸水平,草鱼幼鱼体增重则不再有显著变化。饲料效率和蛋白质积累率表现出与体增重相同的趋势。饲料亮氨酸水平对草鱼幼鱼成活率、脏体比、肝体比和肥满度无显著影响(P>0.05)。
2.2 草鱼幼鱼鱼体必需氨基酸、支链氨基酸和总氨基酸含量情况(见表4)
由表4可知,饲料亮氨酸水平对草鱼幼鱼鱼体必需氨基酸、支链氨基酸和总氨基酸均无显著影响(P>0.05)。
2.3 草鱼幼鱼对饲料亮氨酸需求量
以鱼体增重为指标,经折线模型回归分析,求得草鱼幼鱼对饲料亮氨酸需求量为15.19 g/kg干饲料,约占饲料粗蛋白质的4.68%(见图1)。
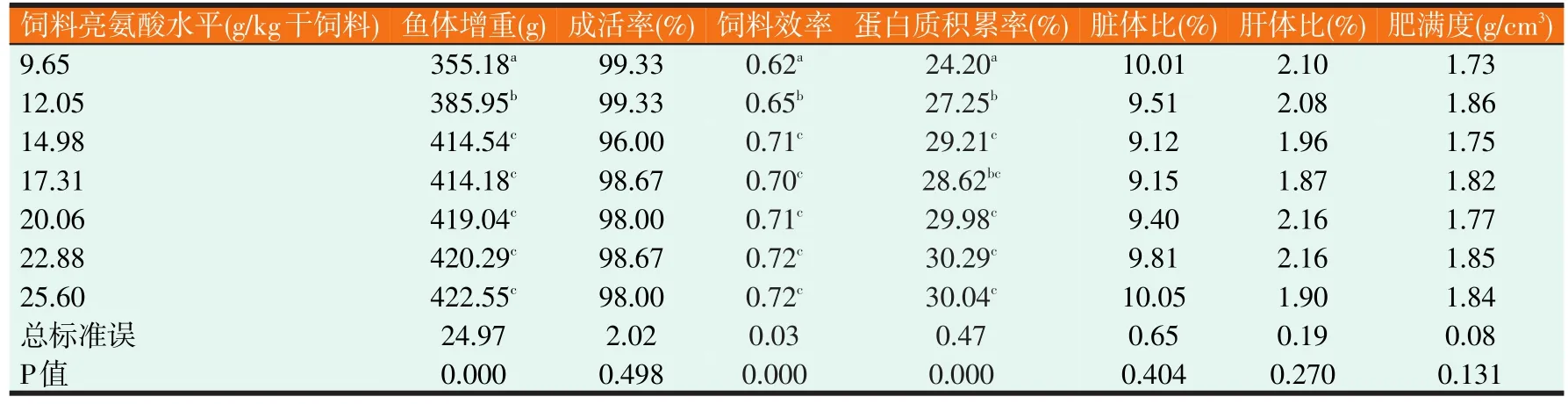
表3 饲料亮氨酸水平对草鱼幼鱼鱼体生长及体形指标的影响
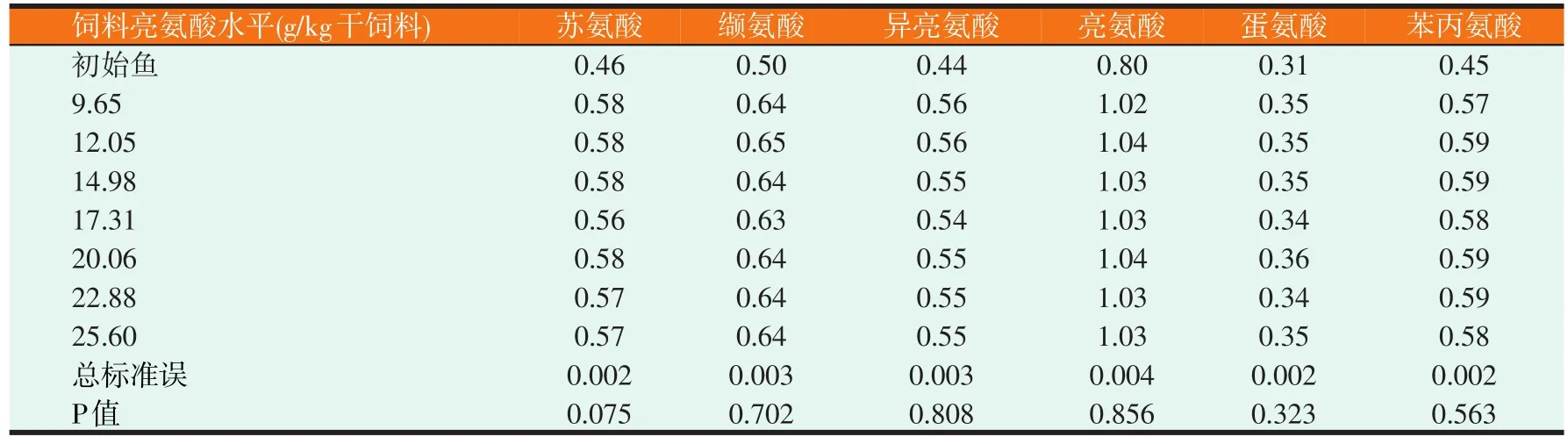
表4 饲料亮氨酸水平对草鱼幼鱼鱼体必需氨基酸、支链氨基酸和总氨基酸含量的影响(%)
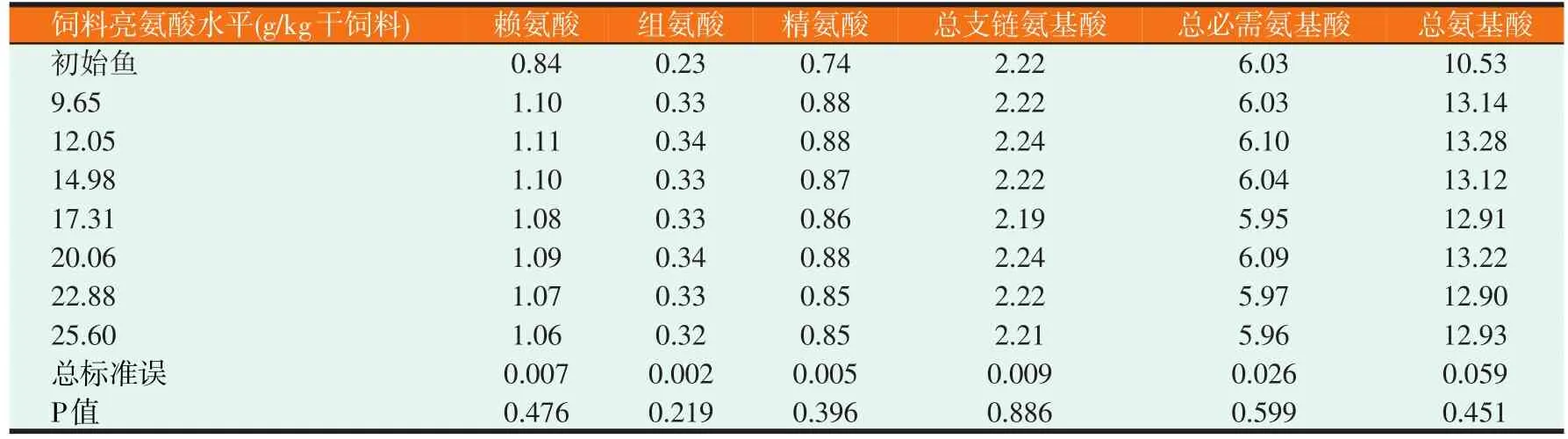
表4 (续) 饲料亮氨酸水平对草鱼幼鱼鱼体必需氨基酸、支链氨基酸和总氨基酸含量的影响(%)

图1 饲料中亮氨酸水平对草鱼幼鱼鱼体增重的影响
以饲料效率为指标,经折线模型回归分析,求得草鱼幼鱼对饲料亮氨酸需求量为15.29 g/kg干饲料,约占饲料粗蛋白质的4.71%(见图2)。
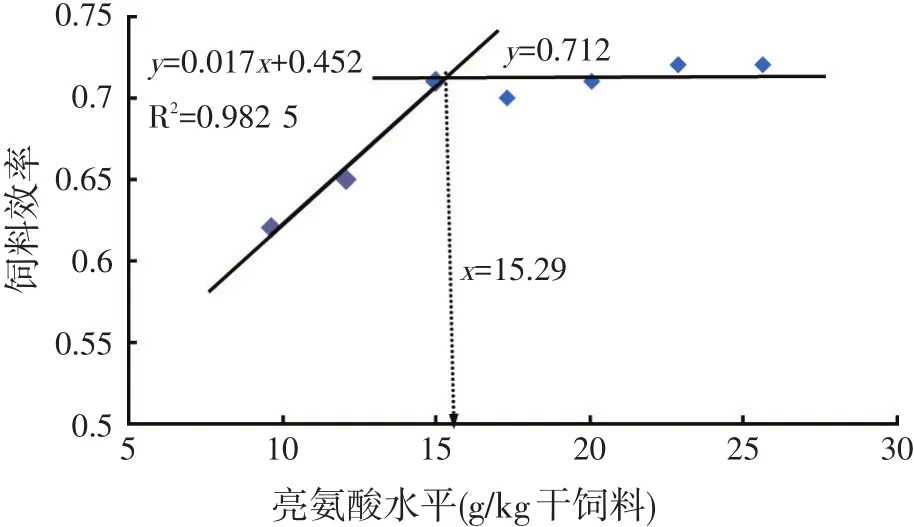
图2 饲料中亮氨酸水平对草鱼幼鱼饲料效率的影响
以蛋白质积累率为指标,经折线模型回归分析,求得草鱼幼鱼对饲料亮氨酸需求量为15.17 g/kg干饲料,约占饲料粗蛋白质的4.68%(见图3)。
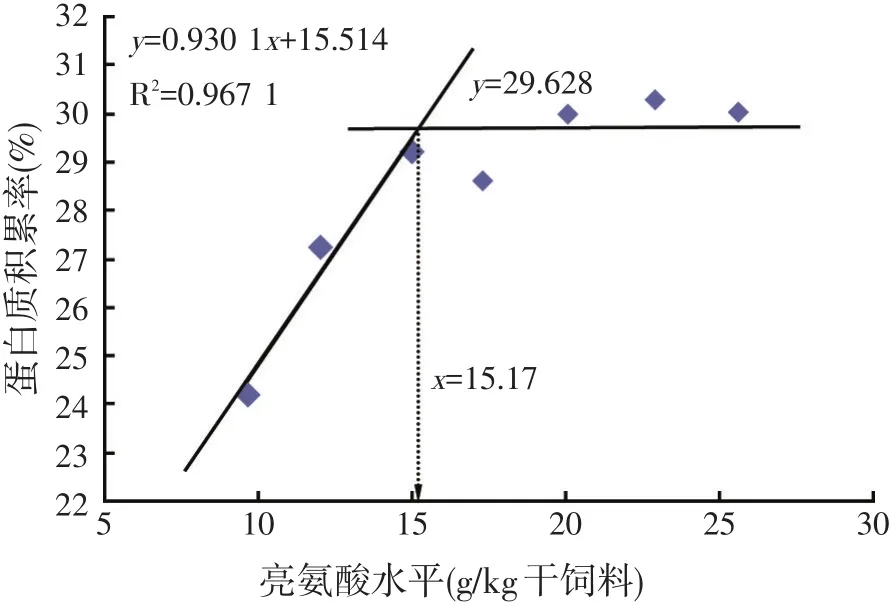
图3 饲料中亮氨酸水平对草鱼幼鱼蛋白质积累率的影响
2.4 草鱼幼鱼全鱼体组成(见表5)
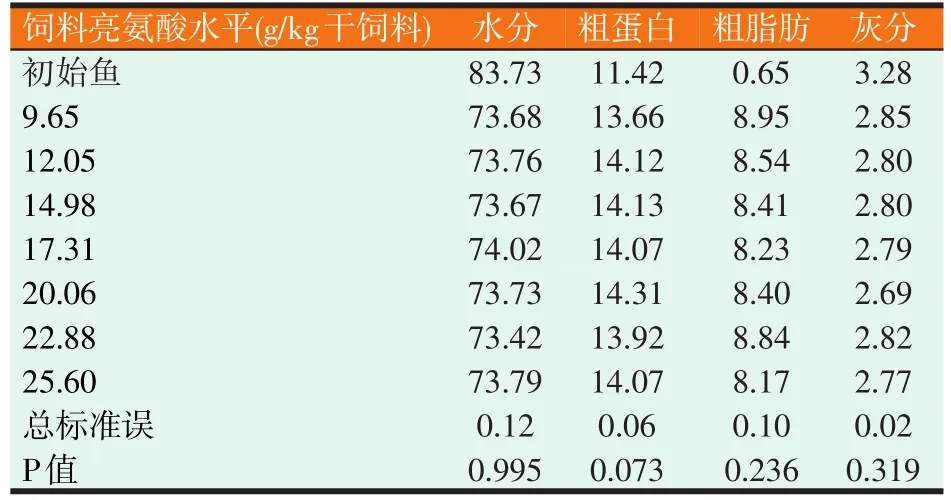
表5 草鱼幼鱼全鱼组成(%)
由表5可知,饲料亮氨酸水平对全鱼水分、粗蛋白质、粗脂肪和灰分无显著影响(P>0.05)。
2.5 草鱼幼鱼肝脏谷丙转氨酶和谷草转氨酶活性(见表6)

表6 草鱼肝脏谷丙转氨酶和谷草转氨酶活性(U/g prot.)
由表6可知,饲料亮氨酸水平对草鱼肝脏谷丙转氨酶和谷草转氨酶活性均无显著影响(P>0.05)。
3 讨论
3.1 饲料中亮氨酸水平对草鱼幼鱼生长、饲料利用和蛋白质积累率的影响
亮氨酸是鱼类的必需氨基酸,在饲料中适当添加亮氨酸可以提高鱼类的生长性能[16]。在本试验中,饲料中亮氨酸含量低于14.98 g/kg干饲料时,随饲料亮氨酸水平降低,草鱼幼鱼生长不断减慢,饲料效率和鱼体蛋白质积累率显著下降。当饲料亮氨酸含量超过14.98 g/kg干饲料时,随饲料中亮氨酸含量升高,草鱼幼鱼生长、饲料效率和鱼体蛋白质积累率则不再发生显著变化。其原因可能是草鱼幼鱼摄食亮氨酸含量低的饲料后,因必需氨基酸不平衡,一方面利用氨基酸合成体蛋白受限,另一方面体内过多的其他氨基酸被分解,使鱼体因排出多余的氨而额外消耗能量,从而影响了草鱼幼鱼生长、饲料利用和蛋白质积累。而当饲料中亮氨酸的含量达到一定水平时,草鱼生长、饲料效率和蛋白质积累率达到最佳,而且进一步提高饲料中亮氨酸的含量,则不再有显著变化。表明此时饲料亮氨酸水平均能满足草鱼正常生长需要,当饲料亮氨酸水平超过最低需要量后,虽然再次打破必需氨基酸平衡,但饲料总氨基酸保持不变,不会给鱼体造成负影响。该结果与对异育银鲫[17]的研究结果相一致,但也有研究者得到的结果与此并不完全一致。对鲈鱼[18-19]、大黄鱼[18]、建鲤[20]、吉富罗非鱼[21]和卵形鲳鲹[22]的研究表明,随着饲料亮氨酸水平的升高,鱼体生长、饲料效率和蛋白质积累率均显著提高,但进一步增加饲料亮氨酸水平,鱼体的生长、饲料效率和蛋白质积累率则显著降低,饲料中过量的亮氨酸抑制鱼类生长主要有两方面原因:一是饲料中过量的亮氨酸会氧化分解生成大量的酮等有毒物质,毒害鱼类机体[23];另一方面是支链氨基酸的颉颃作用,因载体竞争而影响其在小肠壁的吸收和转运[24]。当体内某种支链氨基酸含量过高时,支链氨基酸转氨酶和α-酮酸脱氢酶复合物的活性增加,可增强分解其他两种支链氨基酸,导致这两种支链氨基酸缺乏,破坏氨基酸平衡,降低鱼体氨基酸利用率,抑制鱼体生长[17]。
3.2 草鱼幼鱼对亮氨酸的需要量
鱼类对亮氨酸的需要量主要根据生长实验出现的剂量-生长效应,采用不同数学模型加以确定[25]。已有报道采用较多的数学模型主要有折线模型(Broken-line regression)或二次曲线模型(Quadratic regression)。本试验采用鱼体增重作为效应指标,得出草鱼幼鱼对饲料亮氨酸的需求量为15.19 g/kg。在研究鱼类亮氨酸需求量时,除用生长指标作为确定需求量依据外,许多研究者还用其他的指标加以验证,发现饲料效率、蛋白质利用率、蛋白质积累率、亮氨酸沉积率、肌肉氨基酸含量等也是非常好的有效指标[21-22,26]。本试验分别采用饲料效率和蛋白质积累率作为效应指标得出草鱼幼鱼亮氨酸需求量分别为15.29 g/kg和15.17 g/kg,与生长指标得出的需求量比较相近,表明饲料效率和蛋白质积累率作为确定亮氨酸需求量验证指标也是有效的。因此草鱼幼鱼对饲料亮氨酸适宜需求量为15.17~15.29 g/kg干饲料,占饲料粗蛋白质的4.68%~4.71%。这一结果低于花鲈[18]的5.76%,大黄鱼[18]的6.79%,吉富罗非鱼[21]的7.27%、花鲈[19]的6.54%~6.63%、卵形鲳鲹[22]的7.15%~7.46%,与异育银鲫[17]的4.39%~4.96%相近,高于建鲤[20]的12.9 g/kg干饲料、印度囊鳃鲶[26]的4.34%,造成这种差异的原因可能是试验鱼的种类、大小、饲料的组成及适口性、投喂次数及水平、试验条件等的不同[27-28]。另外,本试验草鱼幼鱼(2.25 g)对饲料亮氨酸需求量占饲料蛋白的4.68%~4.71%高于大规格草鱼[(295.85±2.07)g][14]的4.20%~4.30%,即鱼类规格越小对饲料中氨基酸需求量越高。
3.3 饲料中亮氨酸水平对草鱼幼鱼鱼体必需氨基酸、亮氨酸、异亮氨酸和总氨基酸含量的影响
鱼类的生长主要是对饲料中蛋白质的积累过程,蛋白质的积累又是通过氨基酸的合成来实现的。目前,关于饲料亮氨酸水平对鱼体氨基酸含量的影响已有报道。孙玉轩[21]研究发现饲料亮氨酸水平对吉富罗非鱼肌肉氨基酸组成影响显著。Kim等[29]研究发现饲料亮氨酸水平对牙鲆全鱼氨基酸组成无显著影响。路凯[19]研究发现饲料亮氨酸水平对花鲈肌肉氨基酸组成影响不显著。本试验结果与后两者相同,饲料亮氨酸水平对草鱼幼鱼鱼体各必需氨基酸及其总量、各支链氨基酸及其总量均无显著影响(P>0.05)。出现该结果的原因可能是由于肌肉氨基酸组成相对保守,不受营养水平影响[19]。
3.4 饲料亮氨酸水平对草鱼幼鱼全鱼体组成的影响
国内外学者就饲料亮氨酸水平对鱼体组成的影响已有一些报道。Farhat等[26]研究发现,饲料亮氨酸水平对印度囊鳃鲶[(6.8±0.2)g]全鱼水分、粗蛋白质、粗脂肪和灰分均影响显著。孙玉轩[21]研究发现,饲料亮氨酸水平对吉富罗非鱼[(53.65±0.08)g]全鱼粗蛋白质和粗脂肪影响显著,对全鱼水分则无显著影响。谭小红[22]研究发现饲料亮氨酸水平对卵形鲳鲹[(18.81±0.18)g]全鱼水分、粗蛋白质和粗脂肪影响显著,对灰分则无显著影响。伍曦[20]研究发现,饲料亮氨酸水平对建鲤[(7.88±0.02)g]全鱼水分、粗蛋白质、粗脂肪和灰分均有显著影响。而李燕[18]对鲈鱼[(8.0±0.2)g]和大黄鱼[(6.0±0.1)g]、李桂梅[17]对异育银鲫[(3.15±0.01)g]、路凯[19]对花鲈[(167.82±2.16)g]的研究则表明饲料亮氨酸水平对以上鱼类体组成均无显著影响。本试验结果表明饲料亮氨酸水平对草鱼全鱼水分、粗蛋白质、粗脂肪和灰分均无显著影响。以上研究者得到不同结论的原因除与鱼的种类不同有关外,还可能与试验鱼生长阶段、生长速度、实验剂量范围等因素存在差异有关。
3.5 饲料亮氨酸水平对草鱼肝脏谷丙转氨酶和谷草转氨酶活性的影响
ALT和AST是机体最重要的两种转氨酶,广泛分布于机体的各组织器官中,通常存在于细胞膜、细胞质和线粒体中,这两种酶在机体蛋白质和氨基酸代谢中起重要作用,其活力与氨基酸代谢强弱有关[17]。肝脏是生物体进行各项生命活动的酶活反应场所,故ALT和AST在肝脏中活力较高。伍曦[20]研究发现亮氨酸显著影响幼建鲤肝脏谷丙转氨酶和谷草转氨酶活力。路凯[19]对花鲈的研究和谭小红[22]对卵形鲳鲹的研究得到的结果与此相一致。李桂梅[17]对异育银鲫的研究发现饲料亮氨酸水平显著影响肝脏ALT活力,对AST活力则影响不显著。而本试验中,草鱼幼鱼肝脏ALT和AST活力均不受饲料亮氨酸水平的影响。饲料亮氨酸水平与鱼类肝脏ALT和AST活力的关系有待进一步探讨。
4 结论
本试验条件下,分别以鱼体增重、饲料效率和蛋白质积累率为指标,采用折线模型回归分析,求得草鱼幼鱼对饲料亮氨酸的需求量为15.17~15.29 g/kg干饲料,约占饲料蛋白质的4.68%~4.71%。饲料亮氨酸水平对草鱼幼鱼体组成和氨基酸组成均无显著影响。
猜你喜欢
水电与抽水蓄能(2021年2期)2021-05-14 01:42:26
农业工程学报(2021年4期)2021-05-09 08:23:58
农业工程学报(2020年6期)2020-05-19 07:46:38
动物营养学报(2015年10期)2015-12-01 03:30:54
动物营养学报(2015年10期)2015-12-01 02:26:22
动物营养学报(2015年10期)2015-12-01 02:26:21
应用海洋学学报(2014年1期)2014-11-22 07:17:50
大连工业大学学报(2014年2期)2014-09-19 08:52:52
食品工业科技(2014年9期)2014-03-11 18:15:56
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:21:00
