
光、凋落物和根茎浸提液对剑叶金鸡菊种子萌发和幼苗生长的影响
2018-12-28 05:36曾建军邱洪龙
井冈山大学学报(自然科学版) 2018年5期
曾建军,周 兵,邱洪龙
光、凋落物和根茎浸提液对剑叶金鸡菊种子萌发和幼苗生长的影响
*曾建军,周兵,邱洪龙
(井冈山大学生命科学学院,江西,吉安 343009)
以剑叶金鸡菊种子为材料,分析不同光照强度(100%光强、30%光强、黑暗)和剑叶金鸡菊的凋落物及根茎浸提液3个浓度梯度(0.005、0.025、0.125 g·mL-1)处理下的种子发芽率和幼苗生长差异。结果表明:剑叶金鸡菊种子无论在全光照还是黑暗条件下,发芽率无显著差异。但是与全光照条件相比,30%光照和全黑暗培养条件下对幼苗的生长抑制作用极显著影响。剑叶金鸡菊凋落物和根茎浸提液均极显著抑制种子萌发和幼苗的生长。在种群郁闭作用、凋落物分解物和根茎分泌物的自毒作用综合效应下,剑叶金鸡菊种群内种子萌发和实生苗的生长受到抑制,种内竞争减少。
入侵植物;剑叶金鸡菊;种子;光强;自毒作用
繁殖是外来植物入侵和扩张的重要环节,大多数入侵植物除有性繁殖方式外,克隆繁殖能力也非常强[1]。克隆生长在空间和资源利用方面有着明显的优势,但不足的是克隆后代扩散范围有限[2-4]。相比克隆生长,有性后代扩散的距离远,在拓殖新生境方面要优于克隆后代,因此有性繁殖能力强对大多入侵植物来讲是非常重要的特征[5-7]。然而,具有克隆特性的入侵植物种群中,往往实生苗的更替水平非常低[8],研究影响入侵植物实生苗更替的环境因子,对阐明入侵植物的入侵机理及防治具有十分重要的意义[9]。
剑叶金鸡菊()为菊科金鸡菊属植物,原产于北美,属多年生草本。1936年作为园林植物引入江西庐山[10],为中国主要外来入侵物种[11]。如今,长江流域中下游的大部分区域,已成为为金鸡菊属入侵植物的高适生区[12]。剑叶金鸡菊既可通过有性,也可通过克隆方式进行繁殖,其花果期长达6-7个月,结实量高,每株植株种子产量约为12000粒[13]。虽然剑叶金鸡菊有性繁殖能力非常强,但其单优种群实生苗的更替水平比较低[13]。探讨影响入侵植物种群实生苗更替的影响因素,对揭示入侵植物的入侵机制有重要意义。
植物种子萌发和幼苗生长会受许多环境因素的影响,其中光和植物凋落物是其最主要的影响因素[14-15]。剑叶金鸡菊属多年生密集型克隆生长植物,地下根茎非常发达[16],且只有在全光生境才具有最优种群扩散能力[17]。本研究以剑叶金鸡菊种子为实验材料,通过室内生物测定的方法观测光、剑叶金鸡菊凋落物及根茎浸提液对其种子萌发和幼苗生长的情况,以揭示影响剑叶金鸡菊种群实生苗更替的生态机制。
1 材料与方法
1.1 实验材料
供试剑叶金鸡菊种子分别于实验当年采自于江西省吉安市井冈山大学校园内,室温干燥、储藏备用,挑选籽粒饱满,大小均匀的种子用于实验。
1.2 实验方法
1.2.1光对剑叶金鸡菊种子萌发和幼苗生长的影响
2012年9月按照姜勇等[18]方法:不同光强处理是将医用纱布叠起覆盖在培养皿上,通过逐渐增加纱布层数达到 100 %、30 % 2个水平,其中30 %光强设置参照曾建军等[17]模拟林缘相对光强;黑暗处理培养皿放置暗培养箱中培养。
选取外形、大小和饱满程度较一致的剑叶金鸡菊种子用1 %的高锰酸钾浸泡15 min进行消毒,用蒸馏水反复冲洗干净,然后用温水浸种24 h,并用滤纸吸干种子表面多余水分种子萌发,每皿放剑叶金鸡菊种子各50 粒,4个重复。遮光处理和全光照处理培养箱设置为白天16 h黑暗8 h,培养温度为(25 ± 2)℃。
以子叶展开为正常发芽标准,对种子发芽进行统计,当连续3 d无新发芽种子出现,即视为发芽过程结束[19]。7 d后测定芽长、根长、发芽率。发芽率(%)= 发芽种子数/供试种子数×100%。
使用方差分析光照对发芽指标的影响,使用Duncan′s多重比较方法,对同一因素内不同水平的差异显著性进行检验。
1.2.2凋落物和根茎浸提液对剑叶金鸡菊种子萌发和幼苗生长的影响
剑叶金鸡菊凋落物和根茎浸提液:2014年11月份选取选择自然凋落到地表的干净、新近未分解的剑叶金鸡菊地上部分,并挖取地下根茎部分,洗净晾干,用剪刀剪成细碎片,称取12.5 g,加入100 mL蒸馏水,室温浸提72 h。三层纱布过滤得到浓度为0.125 g·mL-1的浸提液,浸提液加蒸馏水稀释成浓度为0.005、0.025 g·mL-1,保存于4 ℃冰箱中待用。
2015年4月,按照1.2.1方法处理种子,每皿放置50粒,并加入不同浓度供试浸提液15 mL,每个浓度处理重复3次,设蒸馏水作为对照,在25 ℃、80 %湿度、光照16 h的培养箱中培养。培养箱内培养处理1 d后统计各处理种子的发芽数(以芽长1 mm为准),每天统计一次,每天记录发芽种子的数量,7 d后计算发芽率、化感效应敏感指数。
发芽率= (发芽种子总数/供试种子总数)×100%
发芽指数:GI = Σ(Gt/Dt)
式中,Gt为逐日发芽种子数,Dt为相应发芽天数。
活力指数:VI = GI×S
式中,GI为发芽指数,S为第7天测得的整株鲜重(g)。
发芽率化感效应敏感指数(RI):
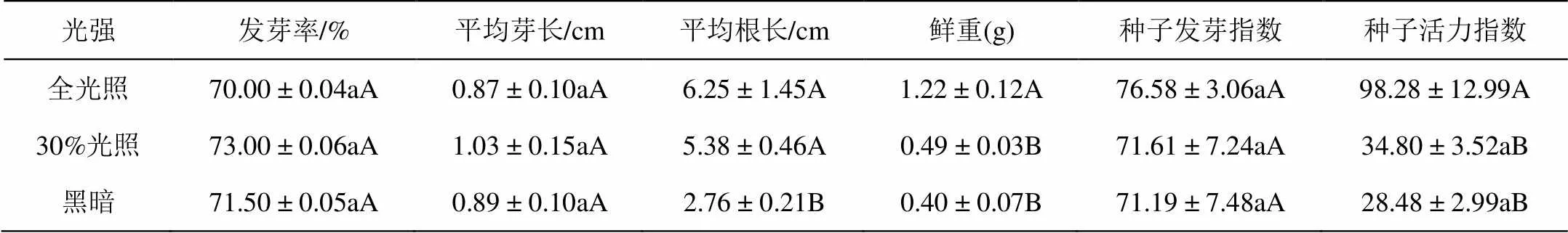
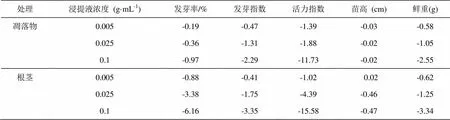
RI = 1-C/T(T≥C),或者RI = C/T-1(T 式中,C为对照值,T为处理值。RI表示化感作用强度大小,正值表示促进效应,负值表示抑制效应,其绝对值大小反映化感作用的强弱[20-21]。 化感综合效应( SE):其是指同一处理下对同一受体各测试项目化感效应指数(RI)的算术平均值[21]。 表1所示,剑叶金鸡菊种子在不同光照条件下,发芽率和种子发芽指数无显著差异。但是与全光照条件相比,30 %光照和全黑暗培养条件下培养7 d,幼苗的生长受到极显著影响,近似林缘30%光照条件下幼苗重量下降了59.83 %。光主要影响根的生长,黑暗条件下根长与对照相比差异达到极显著水平(< 0.01),根长抑制率达到55.84 %。与全关照对比,遮光和黑暗极显著降低种子活力指数(< 0.01)。 表1 不同光强对剑叶金鸡菊种子萌发和幼苗生长的影响 注:同列数据后标不同小写字母表示在5%水平的差异(< 0.05);标不同大写字母表示在1%水平的差异(< 0.01)。下同。 凋落物和根茎浸提液均显著降低剑叶金鸡菊种子萌发率和幼苗的生长。从表2可知,凋落物浸提液浓度为0.005 g·mL-1时,种子的发芽率下降了16.19%,当浓度达到0.125 g·mL-1时,种子的发芽率下降了49.2%。凋落物浸提液对幼苗芽和根的生长的抑制率最高分别达到29.72%和30.22%。与凋落物浸提液比较,根茎浸提液对剑叶金鸡菊种子萌发及幼苗的生长抑制更强烈。根茎浸提液0.005 g·mL-1时,种子萌发率的抑制作用下降了46.66%,当浓度达到0.125 g·mL-1时种子萌发率的抑制作用可下降80.03 %,种子萌发率极低。无论是凋落物还是根茎浸提液,对剑叶金鸡菊种子发芽指数和种子活力指数均有极显著的抑制作用(< 0.01)。 表2 凋落物和根茎浸提液对剑叶金鸡菊对种子萌发和幼苗生长的影响 凋落物和根茎浸提液对剑叶金鸡菊种子萌发及幼苗生长自毒抑制作用较大(表3),地下茎浸提液对种子发芽率、根长、鲜重的抑制作用高于凋落物浸提液,化感指数分别为-6.16、-1.56、-3.34,其中对根的生长抑制作用大于对芽的生长抑制作用。两种浸提液对种子活力指数影响显著,分别为-11.73和-15.58。 表3 凋落物和根茎浸提液对剑叶金鸡菊种子发芽率、发芽指数、活力指数,幼苗的苗高、根长及鲜重的化感效应 由表4知,剑叶金鸡菊凋落物和根茎浸提液对其种子的萌发和幼苗的生长综合效应表现一致,均为抑制作用,且抑制作用与浓度大小呈正相关。在0.005 g·mL-1低浓度下,叶和地下茎浸提液的综合化感效应最低,且两种浸提液的抑制效应一致。随浸提液浓度的升高,抑制作用逐渐增强。0.125 g·mL-1浓度下,叶和根茎浸提液的综合效应分别为-3.00和-5.08,与低浓度相比抑制作用分别增加了6.12倍和10.36倍,与叶浸提液相比,高浓度根茎浸提液的抑制作用更强。 表4 凋落物和根茎浸提液对剑叶金鸡菊种子萌发的综合化感效应 光照不仅影响植物的光合生理,而且影响种子萌发和幼苗的生长[22-23]。许慧男等[19]研究表明,许多入侵植物种子往往是需光发芽,即有光条件下平均高出无光条件发芽率,如入侵植物薇甘菊种子只能在有光的条件下萌发,而在没有光照的条件下萌发率几乎为零。姜勇等[23]研究认为入侵植物紫茎泽兰种子需光才能萌发,种子在土壤中无光环境可形成数量可观的种子库,一旦表土破坏,外界光照能够刺激种子大量萌发,是紫茎泽兰入侵扩散的重要机制之一。不同于薇甘菊和紫茎泽兰,光未影响剑叶金鸡菊种子的萌发,但是光强影响剑叶金鸡菊幼苗的生长。与全光照相比,近似林缘光照条件下(30%光照)幼苗重量下降了59.83%。因此,自然条件下,剑叶金鸡菊种群内及种群边缘虽可见大量实生苗萌发,但只有在全光生境条件下,实生苗才能正常生长。在郁闭度较高的种群内及光照强度较弱的林缘周围,由于光照弱导致剑叶金鸡菊实生苗生长瘦弱甚至死亡。综上,剑叶金鸡菊种子萌发不需光的特性使其种子成熟散发后当年即可萌发,这种特性有利于剑叶金鸡菊占据裸地全光照生境,是其形成入侵的重要机制。 外来入侵植物通过根部和凋落物扩散的化感物质会逐渐在土壤中累积,因此随着入侵植物产生的化感物质浓度不断增加,自毒作用产生,入侵植物自身的重建和生长将受到影响[24-26]。剑叶金鸡菊为多年生根茎类克隆植物,每年秋冬季在基株根蘖处萌发大量密集型克隆分株,同时茎枝倒伏后节间腋芽萌发形成游击型克隆分株,极容易形成单优种群[13, 16]。剑叶金鸡菊单优种群中,每年秋冬季有大量地上部分凋落物,加之根茎分泌物势必会影响其种子的萌发和幼苗的生长,而种子萌发阶段是植物生活周期中最不能忍受不利环境因素的阶段[27-28]。我们的研究表明,不同浓度剑叶金鸡菊凋落物和根茎水浸提液(0.005、0.025、0.1 25 g·mL-1),尤其是多年生根茎浸提液对其自身种子的发芽及幼苗生长有显著的抑制作用,且抑制作用随着浓度增大而增强。因此,在剑叶金鸡菊的单优种群中,种子苗的重建和生长受到抑制。 李愈哲等[29]在研究加拿大一枝黄花的自毒作用时,认为自毒作用使其单优群落可能存在周期性的自发衰退现象。笔者在江西庐山剑叶金鸡菊种群长期定位观察点观察到,其单优种群也存在衰退现象。我们推测,自毒作用是导致剑叶金鸡菊单优群落自发衰退现象的重要原因之一。在庐山,因旅游业发展,剑叶金鸡菊花被大量采摘用于制作花环,有性繁殖受到抑制,剑叶金鸡菊在庐山的扩散明显受到遏制。因此,有性繁殖是剑叶金鸡菊快速扩张的重要途径,阻挡种子传播是防控入侵植物剑叶金鸡菊的有效措施。 [1] Swope S M, Parker I M. Widespread seed limitation affects plant density but not population trajectory in the invasive plant centaurea solstitialis[J]. Oecologia, 2010, 164:117-128. [2] Otfinowski R, Kenkel N C. Clonal integration facilitates the proliferation of smooth brome clones invading northern fescue prairies[J]. Plant Ecology, 2008, 199: 235-242. [3] Liu F, Liao YY, Li W, et al. The effect of pollination on resource allocation among sexual reproduction, clonal reproduction, and vegetative growth in(Alismataceae)[J]. Ecological Research, 2010, 25: 495-499. [4] Roiloa SR, Rodriguez-Echeverria S, delaPena E,et al.Physiological integration increases the survival and growth of the clonal invader Carpobrotus edulis[J]. Biological Invasions, 2010, 12: 1815-1823. [5] Prati D, Schmid B. Genetic differentiation of life-history traits within populations of the clonal plant Ranunculus reptans[J]. Oikos, 2000, 90:442-456. [6] Eckert C G, Lui K, Bronson K, et al. Population genetic consequences of extreme variation in sexual and clonal reproduction in an aquatic plant [J]. Molecular Ecology, 2003,12:331 - 344. [7] Grimsby J L, Tsirelson D, Gammon M A, et al. Genetic diversity and clonal vs. sexual reproduction in Fallopia spp. (Polygonaceae)[J]. American Journal of Botany, 2007, 94:957-964. [8] Harald A, Roldan B. Seedling recruitment in the invasive clonal shrub, Mahonia aquifolium Pursh (Nutt.)[J]. Oecologia1997, 110: 205–211. [9] Qi S S, Dai ZHC, Miao SL, et al. Light limitation and litter of an invasive clonal plant,, inhibit its seedling recruitment[J]. Annals of Botany, 2014, 114: 425-433. [10] 万慧霖,冯宗炜,庞宏东. 中国庐山外来植物[J]. 生态学报, 2008, 28(1):103-110. [11] 徐海根,强胜. 中国外来入侵物种编目[M]. 北京:科学出版社, 2004. [12] 彭冶,王焱,顾慧,等. 外来观赏植物大花金鸡菊在中国的潜在地理分布预测[J]. 南京林业大学学报:自然科学版, 2016, 40(1):53-58. [13] 曾建军,肖宜安,孙敏. 入侵植物剑叶金鸡菊繁殖特征及其与入侵性之间的关系[J]. 植物生态学报,2010, 34(8):966 -972. [14] Sonohat G, Balandier P, Ruchaud F. Predicting solar radiation transmittance in the understory of even-aged coniferous stands in temperate forests[J]. Annals of Forest Science, 2004, 61: 629-641. [15] Valera-Burgos J, Diaz-Barradas M C, Zunzunegui M. Effects of pinus pinea litter on seed germination and seedling performance of three Mediterranean shrub species[J]. Plant Growth Regulation, 2012, 66: 285-292. [16] 曾建军,肖宜安,周小军. 入侵植物剑叶金鸡菊克隆繁殖特性及群落特征[J]. 植物研究, 2012, 32(2):147-150. [17] 曾建军,肖宜安,孙敏. 不同光强下入侵植物剑叶金鸡菊与伴生种生长和光合特征研究[J].广西植物,2011,31(5): 646-650. [18] 姜勇,王文杰,李艳红,等. 光质、光强对入侵植物紫茎泽兰种子萌发及幼苗状态的影响[J]. 植物研究, 2012, 32(4):415-419. [19] 许慧男,王文杰,于兴洋,等. 菊科几种入侵和非入侵植物种子需光发芽特性差异[J]. 生态学报, 2010,30(13): 3433-3440. [20] 慕小倩,何红花,董志刚. 2种杂草水提液对小麦种子萌发及幼苗生长的影响[J]. 西北植物学报, 2008, 28( 6): 1165-1171. [21] 张权,姚小华,滕建华,等. 薄壳山核桃根系水浸提液对 2 种作物种子萌发及幼苗生长的影响[J].植物研究, 2016, 36( 2) :204 -210. [22] 张风娟,李继,徐兴友,等. 环境因子对黄顶菊种子萌发的影响[J]. 生态学报, 2009, 29(4) : 1949-1953. [23] 姜勇,李艳红,王文杰,等.光和不同打破种子休眠方法对紫茎泽兰种子萌发及幼苗状态的影响[J]. 生态学报, 2013,33(1):302-309. [24] Butcko V M, Jensen R J. Evidence of tissue-specific allelopathic activity inand(Asteraceae)[J]. The American midland naturalist, 2002, 148(2):253-262. [25] Wu A P, Yu H, Gao S Q, et al. Differential belowground allelopathic effects of leaf and root of Mikania micrantha[J]. Trees-Structure and Function2009, 23:11-17. [26] Inderjit, Evans H, Crocoll C, et al. Volatile chemicals from leaf litter are associated with invasiveness of a Neotropical weed in Asia[J]. Ecology, 2011, 92:316-324. [27] Kim S, Rayburn A L, Voigt T, et al. Salinity effects on germination and plant growth of prairie cordgrass and Switchgrass[J]. Bioenergy Research, 2012, 5(1):225-235. [28] 徐长林,鱼小军,景媛媛,等. 牦牛粪和藏羊粪浸提液对高寒草甸六种植物种子萌发特性的影响[J].生态学杂志, 2014,33(11):2988-2994. [29] 李愈哲,樊江文,尹昕,等. 入侵植物加拿大一枝黄花与乡土植物芦苇的相互化感作用[J]. 应用生态学报, 2011,22(5):1373-1380. EFFECTS OF LIGHT AND WATER EXTRACTION FROM LITTER AND RHIZOME ON THE SEED GERMINATION AND SEEDLING GROWTH OF THE INVASIVE ALIEN SPECIES OF *ZENG Jian-jun, ZHOU Bing, QIU Hong-long (School of Life Sciences, Jinggangshan University, Ji’an, Jiangxi 343009, China) In the present study, effects of light and water extraction from litter and rhizome on the seed germination and seedling growth of the invasive alien species ofwere studied. The results showed that the light intensity had no significant influence on the parameters of the seed germination, however, the seedling fresh weigh under 30% illumination and darkness was significantly lower than that under 100% illumination. Furthermore, results also revealed that water extraction from litter or rhizome significantly inhibited the seed germination and seedling growth. Under the integrated effects of population closure and the autotoxicity of litter and root exudates, the seed germination and seedling growth were significantly inhibited inpopulation, and the intraspecific competition decreased. invasive plant;; seed; light intensity; autotoxicity 1674-8085(2018)05-0044-06 Q945.34 A 10.3969/j.issn.1674-8085.2018.05.009 2018-06-06; 2018-08-16 国家自然科学基金项目(31760123)资助 *曾建军(1973-),女,江西吉安人,副教授,博士,主要从事植物生态学方向的研究(E-mail: 764178121@qq.com); 周 兵(1977-),男,湖北人黄冈人,副教授,博士,主要从事植物生态学方向的研究(E-mail: 1479322382@qq.com); 邱洪龙(1997-),男,江西南昌人,井冈山大学生命科学学院生物技术专业2014级本科生(E-mail: 758076397@qq.com).2 结果与分析
2.1 光强对剑叶金鸡菊种子萌发和幼苗生长的影响
2.2 凋落物和根茎浸提液对剑叶金鸡菊种子的萌发和幼苗生长的影响

2.3 凋落物和根茎浸提液对剑叶金鸡菊种子萌发的综合化感效应

3 讨论
猜你喜欢
中国水稻科学(2021年6期)2021-11-18
江西农业大学学报(2021年3期)2021-07-22
浙江农业科学(2019年1期)2019-01-25
现代园艺(2017年23期)2018-01-18
红土地(2017年1期)2017-06-05
军营文化天地(2017年2期)2017-03-08
神州民俗(学术版)(2017年1期)2017-02-20
扬子江(2016年1期)2016-05-19
杂草学报(2015年2期)2016-01-04
中国烟草学报(2012年1期)2012-04-09
