
从转化角度谈表观遗传学与变态反应性疾病
2018-12-22 01:15rgTostPhD
中华临床免疫和变态反应杂志 2018年5期
Jörg Tost, PhD
在西方国家,包括IgE相关哮喘在内的变态反应性疾病患病率急速上升。例如,目前严重哮喘患病率达8%,激发试验确诊的食物过敏发病率在澳大利亚高达10%,而美国欧洲国家为2%~4%[1-4]。目前变态反应性疾病已经成为医保系统主要支出项目,预计全球每年由于哮喘支出接近1 000亿美元,而仅在美国由于食物过敏每年支出250亿美元[5-6]。
表观遗传学指“无法用DNA序列改变解释的有丝或减数分裂中基因功能的可遗传改变”,或者正如最近所定义的“染色体区域的结构自适应,从而记录、表明和延续活性改变状态”[7]。表观遗传学决定了哪些基因组区域容易表达发生的改变或可塑性改变从而导致可能疾病倾向的生理状态。近年来表观遗传修饰研究包括DNA甲基化、组蛋白翻译后修饰、核小体占据、sncRNA和lncRNA在变态反应性疾病领域得到了广泛关注。表观遗传学可能的确是解释整个生命中免疫反应高度可塑性的关键,同时表观遗传学调节环境中保护和风险因素对哮喘及变态反应性疾病进程的影响[8]。
尽管仍然存在争议,但仍有大量关于变态反应性疾病发病率升高的解释,其中包括卫生学假说、对微生物的不充分暴露和西方饮食特点等[8]。值得注意的是,这些因素可通过表观遗传密码改变而影响细胞自身稳态。本综述简要论述应用表观遗传修饰对哮喘和变态反应性疾病,包括DNA甲基化、组蛋白修饰和sncRNAs(microRNAs[miRNAs])的研究(图1)。
1 DNA甲基化
至少在哮喘患者中,完成了一些表观基因组关联分析研究(EWASs)并得到鉴定,共同主题包括嗜酸性粒细胞和调节性T细胞(Treg)的重要性和这些细胞可能的表观遗传状态改变[9-11]。大规模EWASs发现疾病相关的环境因素,例如产前母亲吸烟或产前产后环境污染,导致与哮喘和变态反应性疾病相关的基因DNA甲基化改变[12-15]。尽管大量研究集中在对血液中免疫细胞的分析(Potaczek等的综述[8]),仍有研究表明,来源于哮喘患者的鼻或小支气管上皮细胞与健康对照存在不同的甲基化水平,并且甲基化水平的差异超过了在血液细胞中的观察所见[16-17]。因此,鼻上皮细胞可作为上呼吸道上皮细胞的良好替代物[18]。然而,EWASs鉴定的DNA甲基化改变的生物学解释和临床意义仍存在一些争议,例如关于所要分析的组织和细胞类型选择问题,多个研究之间缺乏重复性发现的微
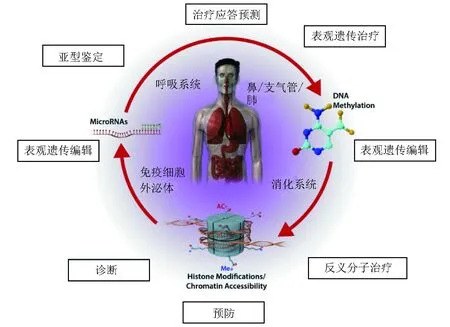
图 1 将表观遗传修饰转化应用于哮喘和变态反应性疾病患者示意图
内圈为靶器官和细胞;正文中详细介绍了不同表观遗传修饰的相互作用(DNA甲基化、组蛋白修饰和小非编码RNAs[miRNAs])和潜在应用价值
小改变对相关功能的影响。此外,大多数研究中对人甲基化组的覆盖率有限,利用表观遗传芯片技术(最常用的全基因组DNA甲基化分析工具)仅仅能覆盖目前已知全部2 900万个CpGs中的450 000到840 000个CpG位点,覆盖率仅为全CpGs的1.6%~2.9%。最大的争议点是这些DNA甲基化变化分析仅反映了特殊的细胞类型比例的变化,但是近期研究不仅发现特殊细胞类型比例变化,同时可以发现特定细胞类型的表观遗传学改变[10,19]。
尽管如此,EWASs具有很大的潜力来解释表型变化,例如在一项DNA甲基化水平联合血清IgE水平的研究中发现,利用全基因DNA甲基化假说来解释IgE浓度的差异优于遗传变异的10倍[20]。EWASs不仅可以通过发现疾病相关通路加深我们对疾病发病机制的了解,如TH1/TH2通路和其他牛奶过敏患者中免疫相关通路,而且可以给未来功能研究提供多个靶点基因,包括转录因子、线粒体蛋白、T细胞突变或氧化应激相关蛋白。尽管有共同目标,但是目前不同的研究均缺乏一致性,这可能由于研究人群、对表型的定义或统计分析方法不同造成的。某些研究发现,不同甲基化位点的特征在出生时已经存在并能预测未来变态反应性疾病的发生[23]。然而,在近期一项关于儿童哮喘的大型队列研究中未发现新生儿时期哮喘相关CpGs甲基化存在差异,提示出生后的环境因素可能是疾病的重要诱因。因此,研究人群的异质性和未知混杂因素可能是限制队列研究结果在出生队列研究中重现的原因[10]。
2 DNA甲基化改变可能作为变态反应性疾病的生物标志物
不同变态反应性疾病在具有同一种上皮屏障的器官中没有表现出同质性,反而形成不同分子表型的高度异质性疾病群组。通过分析炎症类型对患者进行分型并且有利于变态反应性疾病个性化管理的理念,然而,目前个性化治疗缺乏一个强有力且易测的合适核酸类生物标志物。表观遗传变化可能是一个发现、管理甚至预防变态反应性疾病的诊断工具,是评价免疫治疗后耐受和预测早期免疫治疗成功的工具,甚至可用于干预疾病相关通路。
大量的通过血细胞、免疫细胞和上皮细胞分析研究发现,变应性哮喘患者与健康对照者间存在差异[8,16-17]。变态反应性疾病EWASs研究提示,无论是否是易发生变态反应的季节,DNA甲基化特点可以区分变态反应性疾病患者(季节性变应性鼻炎),较基因表达特点鉴别能力更好[24]。通过对脐带血样本92-CpG特点进行分析,可在临床上发现食物过敏之前分析食物过敏进程[23]。通过大规模队列研究证实这些DNA甲基化变化将能用于变态反应性疾病,以早期发现疾病儿童并早期进行免疫治疗干预。
此外,口服食物激发试验是一种耗时、资源集中并且具有风险的诊断方法,目前已提议DNA甲基化特征可作为口服食物激发试验的替代诊断方法,它们还可以帮助变应原测试进行简化和标准化,因为目前的各种化验方法定量方式不同,甚至结果也不一致。一项含有96个CpGs位点的DNA甲基化特征研究显示,其可预示多种食物过敏,并且优于一般的皮肤点刺实验和变应原特异性IgE检测,提示DNA甲基化具有正确诊断变态反应性疾病表观遗传修饰的潜力[25],一项重复队列研究中发现这种方法的诊断准确率接近80%[25]。然而,该研究结果仍需更大规模的独立队列重复研究,这样才能将这些发现转化为临床实践。
3 DNA甲基化治疗和免疫治疗的生物标志物
尽管过去10年中相关研究取得了重大进展,但是仍急需寻求针对哮喘和变态反应性疾病的新型药物和治疗手段。虽然大多数哮喘症状通过常规治疗可控制到稳定状态,但仍有病情严重甚至无法控制的哮喘患者需要两盒其他治疗方法,如单克隆抗体生物制剂。尽管生物制剂如抗IgE抗体奥马珠单抗已被证实能有效控制成人和儿童的严重哮喘症状,并可降低食物过敏口服免疫治疗过程中不良事件的发生频率和严重程度[26-27],但是这些针对IgE、白细胞介素(interleukin,IL)-5、IL-4、IL-13或胸腺基质淋巴细胞生成素的新型治疗被批准用于临床或研发成本对于公共卫生系统来说将是重大挑战[28]。虽然某些生物标志物与治疗反应相关,但是基于分子改变的变态反应性疾病分型更为重要,有助于特殊患者类型的患者对有效药物的选择,改善疗效,同时促进变态反应性疾病患者精准用药。尽管目前的证据局限于癌症领域,如MGMT启动子甲基化作为预测标志及烷基化药物替莫唑胺用于治疗胶质母细胞瘤患者,但是对表观遗传修饰的分析可能有助于更好地定义和了解疾病及其潜在的分子机制,对患者进一步分型,进而选择新的治疗方案,从而最大程度争取有效治疗的机会,同时维持卫生医疗系统的成本。同时,表观遗传分析也可能有助于发现哪类儿童能获得更好的疗效。
目前仍缺乏儿童对于生物制剂疗效的统计数据且有些数据之间存在矛盾结果。近期一项MEDALL财团的多项队列研究发现,儿童时期哮喘与某些甲基化CpG位点不同有关,尤其是血液和嗜酸性粒细胞[10]。对一个亚组纯化后的循环嗜酸性粒细胞分析发现有DNA甲基化改变,提示这是一种不同的活化状态。这些发现为利用抗IgE、抗IL-13或抗IL-5受体拮抗剂抗体进行嗜酸性粒细胞靶向治疗如何改变嗜碱性粒细胞DNA甲基化特点及其疗效与治疗前表观遗传特点是否有联系提供了有趣的依据[30-31]。
全身应用糖皮质激素是目前治疗哮喘的标准方法。初步研究证据表明,DNA甲基化改变可能影响疗效,基因甲基化改变,包括OTX2和VVN1启动子,对于鼻上皮细胞有更好的疗效[32-33]。然而,与基因表达特征不同的是,目前尚未发现DNA甲基化是对糖皮质激素反应程度进行预测的生物标志物[34-35]。尽管如此,这些DNA甲基化标记仍可作为对糖皮质激素不应答的难控制性哮喘患者的辅助判断指标。
在食物过敏领域,诱导和维持对变应原的耐受需要变应原特异性Treg细胞产生。叉头蛋白3(fork head protein 3,FOXP3)调节性T细胞特异性去甲基区(T-cell specific demethylated region,TSDR)的去甲基化是Treg细胞维持抑制作用的先决条件[36-37]。去甲基化有免疫治疗诱导并在对变应原的持续无应答中维持低水平甲基化,如花生或牛奶免疫疗法[38-39]。因此FOXP3去甲基化是免疫疗法是否成功的先决条件。虽然两项研究中样本量较低,但是TSDR对DNA甲基化分析仍有望作为监视免疫治疗是否应答和耐受是否形成的生物标志物。
我们最近在关于花生过敏皮下免疫治疗的小鼠模型中发现,皮下免疫治疗成功小鼠FOXP3甲基化降低,然而,脾CD4+IL-4+T细胞中TH2关键转录因子Gata3显著增高[19]。与此相反,口服免疫治疗仅引起FOXP3去甲基化而无Gata3甲基化,提示后者对于维持无应答水平和防止在皮下免疫治疗中发现的对第二种变应原敏感。
此外,在花生口服免疫治疗中发现了诱导分化出的新的CD4+T细胞亚群[40],尽管到目前为止仅仅是转录水平的特征,但是这些新型CD4+T细胞亚群很可能具有某种特定表观特征,作为与持续无应答相关的标志物。
4 变态反应性疾病患者的组蛋白密码
乙酰化和去乙酰化之间的平衡状态与组蛋白N端残基甲基化和去甲基化一样对调节和控制基因表达发挥重要作用,组蛋白是染色体的重要组构和DNA的基本包装单位,多种组蛋白修饰(所谓的组蛋白密码)和DNA甲基化之间的相互作用决定了转录机器对特定基因组元素的易接近性,并决定特定的细胞系,包括分化为T细胞系(图2)[8,41-42]。尽管相对于DNA甲基化分析相关研究较少,但是在变态反应性疾病和哮喘患者中已发现组蛋白修饰平衡状态的异常,而组蛋白修饰变化会导致染色体螺旋结构的压缩或打开[8,43-44]。值得注意的是,研究已发现来源于哮喘患者T细胞亚群的TH2发育相关基因调节区和哮喘易感基因座的增强子相关组蛋白标志物H3K4Me2增多[45]。在一项母系鼻暴露鲁氏不动杆菌对后代变应性气道炎症起保护效应的小鼠模型中发现是γ干扰素(interferon-γ,IFN-γ)依赖的,至少在部分个体中时通过鲁氏不动杆菌诱导的后代小鼠CD4+T细胞Ifng启动子组蛋白H4乙酰化保护介导的,然而,DNA甲基化分析未发现差异[46]。
大多数组蛋白修饰的分析研究集中于特定基因层面,尤其是TH细胞亚型的分化,很少有系统性的全基因组研究[43]。炎症通常伴随免疫相关基因总而言之,除了CpG富集区(CpG岛)和三分之二的启动子区,其余基因组的CpG二核苷酸都是甲基化状态(带线的实心圆)。在常染色质中,RNA聚合酶Ⅱ能接近启动子CpG岛(带线的空心圆,相当于无核小体区),从这个区域启动转录,而组蛋白是高度乙酰化状态(圈代表乙酰基),组蛋白H3的9号赖氨酸(H3K9)和H3的4号赖氨酸(H3K4)是甲基化状态(多边形代表甲基)。与之相反的,在异染色质中,DNA和核小体高度浓缩,H3K4是去甲基化状态,H3K9、H3K27和H4K20是甲基化状态,DNA也是超甲基话状态,该区域无法转录。为阐明原因,只显示与染色质状态相关的组蛋白修饰。miRNAs以3端非翻译区作为靶点,在完全互补情况下导致靶RNA降解,但大多数情况下,仅有几个碱基配对而导致mRNA无法翻译为蛋白,而不改变mRNA丰度的转录上调和组蛋白的乙酰化,这已在哮喘患者支气管活检标本和肺泡巨噬细胞中得到证实[47]。除组蛋白外,组蛋白乙酰转移酶和组蛋白去乙酰化酶(histone deacetylase,HDACs)还作用于其他一些蛋白,包括变态反应性疾病相关转录因子,例如GATA-3、FOXP3、糖皮质激素受体和核因子κB。研究发现,重度哮喘患者HDACsIII水平降低,导致GATA-3的高乙酰化水平而TH2细胞因子的表达,从而出现气道炎症,小鼠模型中可以利用HDACs拮抗剂[48]或强制表达其他去乙酰化酶家族成员[49]来逆转这种炎性反应。虽然变态反应性疾病患者尚未进行其他类型组蛋白修饰研究,但是有证据表明核染色质修饰酶在气道上皮细胞中发生了改变[50]。
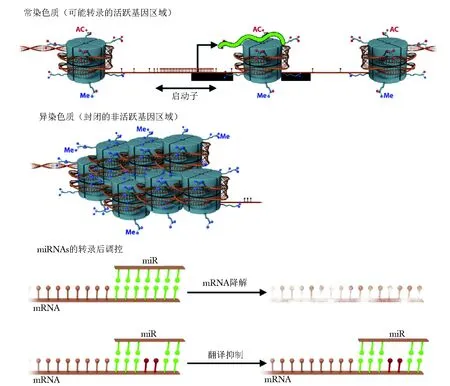
图 2 与开放(常)和闭合(异)染色质相关的表观遗传修饰
5 甲基化组和组蛋白密码作为变态反应性疾病的治疗靶点
表观遗传改变构成了可遗传同时又可逆的基因表达模式,并可以通过调节表观基因组而降低患特应性疾病的风险,或者至少影响不明显表型的疾病进程。非特异性DNA甲基化抑制剂,如5-氮杂胞苷,在变态反应性疾病中尚未研究并且目前的数据存在矛盾。某些研究发现其治疗有效,尤其是可诱导Treg细胞[51]和IFNG去甲基化[52],然而,在研究中会导致过敏症状的加重[53]。作为哮喘的标准抗炎疗法,糖皮质激素可激活几个HDACs,并可取代糖皮质激素受体结合位点的核因子κB,阻断组蛋白乙酰化过程[54-55]。合成糖皮质激素敏感度的下降与HDAC2水平下降有关,HDAC2导致糖皮质激素受体的去乙酰化,被动吸烟可能使其水平下降[56-57]。因此,治疗干预过程中增加HDAC水平有望获得最好疗效。
另一方面,HDAC抑制剂的应用出现了矛盾结果,一些研究表现出抗炎作用而另一些研究提示增强炎症反应,因此这种治疗方法的结果需要进一步研究[44,58-59]。然而,由于HDACs和组蛋白乙酰转移酶作用于大量靶位并参与多种细胞信号通路,所以抑制或调解可能引起意想不到的不良反应,需要研发以特异性细胞亚群为靶点的选择性HDAC抑制剂。
因为大多数研究只针对表观遗传疗法对单个靶位基因的影响,所以对这些非特异的表观遗传修饰的应用效果难以定论,但是有必要对这些疗法保持关注。目前随着基因编辑重大技术的进步,利用crispr/dcas9系统可以对表观基因组进行改造[60-62]。这些方法使我们首次能够在功能上对基因组中任何位点的表观遗传修饰的重要性进行研究和验证,并能提供新的替代方法来调节变态反应性疾病的表观基因组。相比于核酸酶缺陷Cas9酶,CRISPR/dCas9的结合不会引起基因组双链的断裂,从而使我们将表观遗传酶引导到基因组的特定位点上,进而引起特定DNA和组蛋白的甲基化或去甲基化及组蛋白的去乙酰化或乙酰化[63-64]。
尽管这一技术最近才出现,但它在未来对没有明确突变的疾病治疗有着巨大的潜力。因为表观遗传修饰往往是可逆的,所以对环境暴露所引起的关键分子变化的识别可能提供新的治疗方案,以便在出现变态反应症状之前逆转这种变化。更重要的是,变态反应性疾病患者中观察到的T细胞极化主要是由启动子的表观遗传所修饰,由关键转录因子、细胞因子结合位点和保守的非编码序列所驱动。
表观遗传编辑提供了改变不同T细胞亚群平衡的可能工具,这可以通过两个载体来实现,每个载体包含一种细胞类型特异性启动子驱动表达的关键编辑复合体组分,因此导致编辑复合体只在T细胞表达[65]。然而,尽管表观基因组的靶向改造是研究表观遗传变化功能的一个重要进展,并且已在体内进行了第一次实验,但是用于治疗目的的潜在用途仍不确定,其体内应用的安全性和有效性仍不明确。因此,在临床上用于变态反应性疾病的治疗前还需进一步研究其变化效率、诱导变化的稳定性和诱发的大量非特异改变(脱靶效应)[67]。
6 微生物组和表观遗传组的相互作用
微生物通过多种机制影响细胞稳态、免疫细胞分化和极化,包括表观基因组调控[68]。大量研究已经报道了变态反应性疾病患者皮肤、上气道和肠等屏障组织中微生物多样性的变化和普遍性[69-71]。由于目前发表的研究结果对发现的变化和不同种类普及度的鉴定不一致,并且微生物多样性的增加和减少都与变态反应性疾病有关,因此许多问题仍待进一步研究。
变态反应性疾病患者中某些细菌种属发生了改变,例如梭状芽孢杆菌属。这些种属可以产生表观活性免疫调节分子,导致肠道巨噬细胞超乙酰化和TH1/TH17效应T细胞极化,例如来自膳食纤维的短链脂肪酸(short chain fatty acids,SCFAs)包括可以抑制HDACs的丁酸[72-74]。低SCFAs水平与变态反应性疾病有关,SCFAs水平升高可改善上皮屏障功能,减轻病症[73]。丁酸可以抑制2型固有淋巴样细胞增殖和减少TH2型细胞因子和GATA-3表达,在小鼠的变态反应性炎症模型中,使用一种泛HDAC抑制剂可以对这些结果进行重现[75]。值得注意的是,梭状芽孢杆菌产生的SCFAs与食物过敏由关,可通过基因座的组蛋白H3乙酰诱导形成肠Treg细胞,导致TSDR选择性去乙酰化,从而降低对食物变应原敏感性而形成耐受基因环境[77-78]。同样地,双歧杆菌产生的乙酸盐可导致FOXP3启动子超乙酰化[72]。此外,微生物定植通过肠Treg细胞Uhrf1引起DNA甲基化[79]。总之,这些结果提示,微生物表观活性代谢物的修饰可能改变变态反应性疾病患者的疾病状态。
对具有患变态反应性疾病风险的患者进行干预的主要目的是预防疾病[80]。尽管某些保护性的环境和饮食因素是有益的,仍然需要大量研究确定与诱导耐受相关的表观遗传变化。暴露在益生菌或致病细菌中会导致未成熟的肠上皮细胞的暴露特异性DNA甲基化变化[81]。已证实一些益生菌可以增加T细胞反应的表观遗传可塑性,并可能有助于防止对变应原敏感或降低变态反应的发生[39,46]。其他饮食,如多不饱和脂肪酸已表现出有益的干预效果[82-83],并且至少部分调节作用是通过表观遗传改变实现的或与其相关[84-86]。然而,迄今为止,表观遗传分析仅存在小规模地队列研究,需要进一步的机制研究来阐明多不饱和脂肪酸影响表观遗传修饰的方式和方法。
7 miRNAs和胞外膜泡
在不改变基因序列的情况下,miRNA是又一种表观遗传调节机制,影响基因表达但不改变基因组序列,miRNA(22-25个核苷酸)是通过结合mRNA的3’非翻译区域封闭mRNA翻译或改变其稳定性而在转录后调控基因表达的小非编码RNA分子(图2)[87]。每个miRNA可以直接或间接调节多个靶基因,而且每个靶基因也可以受多个miRNAs调节,形成一个复杂的基因调节网络。miRNAs在免疫细胞的发育、分化、突变和活化以及气道重塑、过敏或炎症疾病中起重要作用[88]。与DNA甲基化模式类似,有强有力的证据表明变态反应性疾病患者组织、细胞或体液中miRNA表达特征与健康对照者间存在差异[89-93]。变应原激发可以导致血细胞miRNA表达进一步改变[94],并且至少在嗜酸细胞性食管炎中糖皮质激素可以逆转miRNA失调[95]。
此外,一些miRNA的表达可用于区分不同哮喘亚型,并且与疾病的严重程度相关联,将来有望用于风险评估[91,96-97]。然而,miRNAs的细胞特异性、不同检测技术、不同实验动物模型和患者、异质性表型的小样本量和大多试验缺乏重复队列导致很难从这些数据中得出明确的结论,并在临床上推进基于miRNA的分析和诊断。
尽管由于其低侵入性而促进了其临床研究,但对循环miRNAs的分析却由于肺泡灌洗液、血清、血浆或其他来源,例如呼出气体miRNAs的低浓度而受阻。胞外膜泡(extracellular vesicles,EVs)是一种异质的小膜包囊泡,包括外泌体和含RNA、蛋白、代谢分子和小调节RNAs(例如miRNAs)的细胞微泡,构成了真核细胞细胞间通信的重要组成部分(图3)[98]。不同于父母细胞类型,EVs的内容物是由细胞因子环境决定的,并来自于原始细胞内容的活化选择(图3)。EVs随后通过受体细胞的内吞作用被吸收,与原始细胞的距离相隔甚远。这种摄取过程一定程度上是由EVs表面的蛋白质所决定的,释放物在受体细胞中可以以级联方式刺激或抑制固有和特异性免疫应答[99-100]。
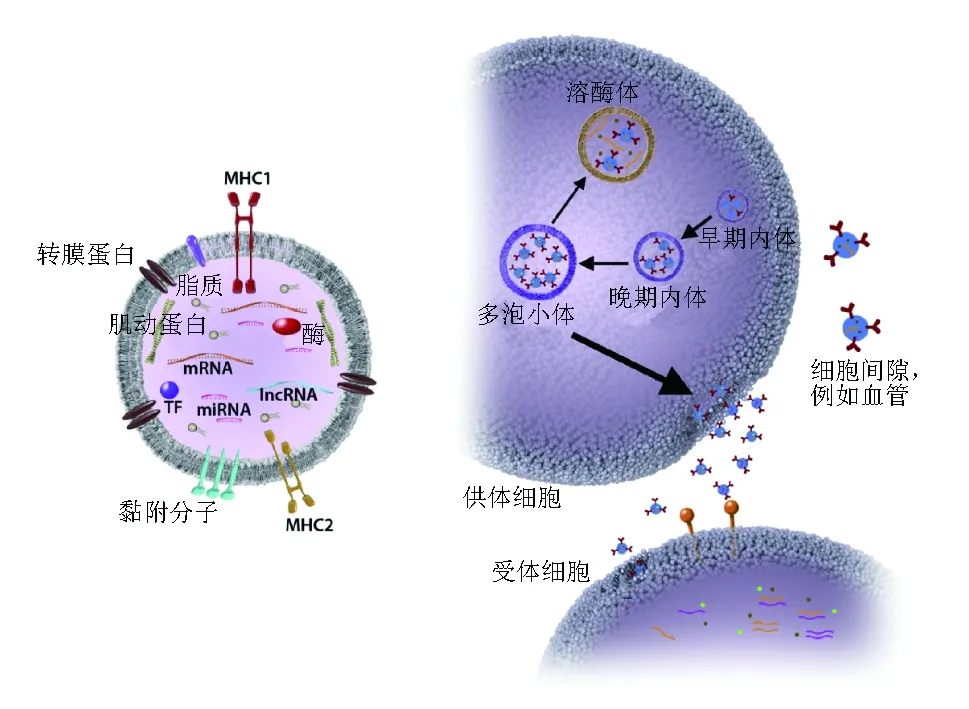
图 3 外泌体内容物及其细胞内信号机制左图显示外泌体包含大量蛋白、脂质和核酸分子,包括细胞骨架蛋白(肌动蛋白和微管蛋白)、酶(甘油-3-磷酸脱氢酶和延长因子)、部分转运所需的内体分选复合物(ESCRT)、转录因子(TF)、信号传导相关蛋白、miRNAs及其他小非编码RNAs、mRNAs和长非编码RNAs(lncRNAs)。在双层脂质膜上具有抗原提呈MHC1和MHC2,细胞黏黏附相关分子(四分子交联体家族和整合素)和其他跨膜蛋白。右图显示小囊泡通过内体内膜出芽方式的产生和吸收过程,包括ESCRT、四分子交联体家族和脂质形成多泡小体,随后与脂质膜融合而释放外泌体。外泌体内容物通过质膜融合或内吞作用利用特定配体受体相互作用被受体细胞所接收,供体和受体细胞在空间上可以存在极大距离
因此外泌体的内容物对受体细胞具有编辑功能并且在过敏调节细胞相互作用中发挥重要作用,利用这些内容物可减轻或加重变态反应[93,103]。变应原致敏导致EVs数量明显增多[104],在一个支气管哮喘小鼠模型中导致TH2抑制性miRNAs选择性进入EVs,从而减少嗜酸性粒细胞积累和气道炎症[105]。利用分离至肺泡灌洗液EVs中的miRNA对鉴别轻度哮喘患者的预测率为72%,包含多种调节疾病相关细胞因子的miRNAs[106],并且与哮喘患者气道上皮细胞失调的miRNAs存在极大重合度[107]。
少数研究分析了miRNAs与变态反应性疾病治疗的关系。miR-21是一种已知的变态反应性疾病患者的miRNA,与对吸入性糖皮质激素(inhaled glucocorticoids,IGCSs)敏感的儿童相比,耐受IGCSs的儿童miR-21水平较高。然而,因为耐药儿童miR-21水平与不适用IGCSs患者的水平接近,故IGCS敏感患者miR-21水平下降可能是由于其哮喘状况的改善而不是对治疗的效果。同理,miRNA的变化与口服糖皮质激素或抗白三烯治疗有关[97]。
8 miRNAs作为变态反应性疾病的靶点
miRNAs提供了一个有意思的干预措施,利用合成miRNA类似物或miRNA抑制剂调节疾病相关的内源性miRNAs的表达。从变态反应性疾病小鼠模型中已经获得了一些研究资料。let7-a(靶点是TH2型细胞因子IL-13)的外源注射可减少细胞因子水平和气道炎症[109]。卵清蛋白(ovalbumin,OVA)诱导的小鼠哮喘模型中miR-221的抑制同样减少嗜酸性粒细胞和白细胞气道浸润[110],尽管效果较差,但是尘螨处理小鼠miR-145的抑制减少了Th2细胞IL-5和IL-13水平,类似糖皮质激素的治疗效果[111]。
由于miRNAs的普遍存在和多重靶点,治疗制剂需要通过细胞类型特异性方式传递,并且预测脱靶率需要对靶点和细胞因子产生的复杂机制有更深一步的了解。此外,由于感兴趣的转录本可以被多个miRNAs调节,因此单个miRNAs水平上进行调制效果有限。
尽管还未用于变态反应性疾病的治疗,但目前已设计了特异性治疗和载体并正在进行临床试验研究,增加(miR-34、mir-29、miR-16)或减少(miR-122、miR-103/107)特异性miRNA水平[112-114]。值得注意的是,一项对变态反应性疾病的临床研究(NCT02580552)以皮肤T细胞淋巴瘤中的miR-155为靶点,因为miR-155特异表达于Th2 CD4+细胞,并且miR-155缺陷鼠哮喘较轻[115-116]。随着基于miRNA的治疗在其他领域的进展,尤其是癌症,细胞类型特异性传递将不断提及,这种方法可能会对哮喘和变态反应性疾病的治疗进行评估。
此外,lncRNAs和环状RNAs已经被证明是miRNA的强抑制物,可减少细胞中特异性miRNAs的水平[117-118]。尽管在生理条件下miRNA水平会适应这种消耗,合成并设计能清除疾病相关的miRNAs将成为潜在的治疗方案[119]。然而,它们的发展和体内传输的效应方法已经远远落后于miRNA类似物和抑制剂了。
在没有天然小RNA的情况下,针对治疗变态反应性疾病的转录靶点,另一个选择是调节细胞分化,这在免疫系统中起重要作用(例如T细胞)。例如,可以通过减少促进TH2应答的转录因子(GATA-3)表达来实现,可以通过反义策略来实现。反义分子通过特定的结合来阻止mRNA转化为蛋白质,随后使mRNAs降解[15]。
最近关于HGD40/SB010(脱氧核酶型反义分子抗TH2重要转录因子GATA3)的研究支持了反义方法可以用来编辑发育T细胞表型的可能性[120-121]。脱氧核酶是具有内部催化活性的反义分子,可在不需要具有酶活性的辅助分子的情况下进行特定的结合,使mRNA失活[15]。1期临床试验证实HGD40/SB010是安全的及可耐受的[120],Ⅱ期随机安慰剂对照多中心临床试验发现其可显著减弱过敏变应性哮喘患者早期和后期临床哮喘症状[121]。
初步数据也显示了EVs在临床方面的应用潜力。骨髓来源的肥大细胞外泌体在小鼠模型中已被证实可减少循环IgE水平,抑制OVA引起的哮喘中肥大细胞的活化,降低炎症反应强度[122]。利用EVs表达的免疫抑制或刺激细胞因子、其他变态反应性疾病相关分子在体内靶向作用于免疫细胞,或使用定制性临床级EVs靶向作用于免疫细胞治疗干预可能成为新的治疗免疫相关疾病的方法,并在癌症领域作为先驱,从树突细胞中提取的纯化外秘体已经用于临床试验的免疫疗法[123-124],可以假设耐受性EVs注射可以抑制变态反应和超敏反应。通过工程改造EVs传输生物分子或药物通过外泌体表面与细胞类型特异性抗体到特异细胞亚群也获得了广泛关注。然而,重构EVs的复杂性是一项挑战,而且需要更好地理解与加载和分泌有关的细胞过程。短期内进一步描述外泌体亚群来鉴定天然形成的特异性来源EVs较为实际,可以通过富集或清除来治疗干预。
9 哪种表观遗传修饰最适合翻译?
随着分析基因组范围和特定位点的DNA甲基化模式工具的进展,结合生物样品在储存期间的DNA甲基化标记的稳定性,有助于在复杂疾病患者中实施基于DNA甲基化的生物标记,与对其进行的组蛋白修改的分析比较仍存在诸多挑战,包括染色体分离。已经建立了针对小RNAs(包括miRNAs)的成熟和标准化分析技术,同样的基因表达分析可以促进临床试验的实施。所有表观遗传修饰的细胞类型特异性,包括miRNAs,如何完全定义细胞亚群仍然存在困难需要研究。然而,miRNAs的基因调控网络比DNA甲基化或在特定位点上的组蛋白修饰变化复杂得多。如果细胞类型是已知的,而感兴趣的细胞较易分离,那么DNA甲基化就满足了一个重要生物标志物的所有标准[125]。另外,对循环miRNAs或EVs内容物的分析可能会克服这个问题,允许对患者进行重复的最小侵入性取样。
值得注意的另一点是不同水平的表观遗传调控的稳定性与动态性。DNA甲基化对刺激有反应,尽管速度要慢得多,但与组蛋白修饰或miRNAs变化相比,有助于维持表观遗传特征的改变,后者在外部或内部刺激后不久将发生强烈变化,但是,一旦状态稳定下来,变化可能会逐渐消失。因此,如果生物标志物用来预测长期结果或作为可靠的诊断,DNA甲基化可作为标志物选择,而对于快速决策(例如单次注射后治疗反应),miRNA可能更加合适。此外,从干预的角度来看,尽管我们的知识有限,但是利用外源合成类似物和抑制剂调节miRNA水平在临床试验中(尤其是癌症)优于DNA甲基化改变和组蛋白修饰作为靶点。
结论
研究表观遗传学将进一步加深我们对疾病机制的理解,一些表观遗传标记可能作为生物标记物指导治疗变态反应性疾病的个性化管理和临床决策,包括检测、管理、以及理想的预防方法。因为大多数研究结果受小样本队列的限制,只具有较低的统计效力,系统性的包含表观遗传分析,尤其是DNA甲基化和miRNAs,大规模的临床试验将大大提高我们确定与治疗应答相关生物标志物的能力,甚至在治疗开始前预测应答效果。然而,细胞表型是由大量细胞调节机制所决定,包括表观遗传修饰。因此单个DNA甲基化水平或miRNA的变化不可能在临床应用中产生足够的特异性和灵敏度。为了获得具有足够效力的生物标志物模板,将需要类似于目前在癌症患者的个性化管理中涌现的那些多层特征[126]。
此外,维持表观基因组特征在其可塑性限度内的机制冗余性可能难以克服。因此,需要系统生物学方法评估不同水平的表观遗传修饰和基因调控机制的复杂相互作用,并更好地选择治疗靶点。
虽然变态反应性疾病表观遗传学研究仍处于早期阶段,由于认识到复杂疾病中表观遗传学变化的重要性增加和近年来表观遗传修饰分析技术的成熟,故随着表观遗传应用于临床实践,我们可以预测,未来几年知识将大规模扩展。
我们知道什么?
·表观遗传学包括DNA甲基化、组蛋白修饰和小非编码RNAs,是免疫细胞分化和可塑性的重要机制。
·哮喘和变态反应性疾病患者免疫细胞亚群和各类体液的表观遗传修饰特征改变与疾病表型或疾病严重程度相关。
·Treg细胞FOXP3去甲基化与其活化和变应原无应答相关。
·EVs,包括外泌体,其内部分子可以远距离改变细胞表型。
·miRNAs类似物和抑制剂,人工合成反义分子在临床试验中表现出良好效果,虽然大多数研究并不是变态反应性疾病领域。
·宿主表观基因组和微生物组相互作用密切,并且微生物组的调节充分对全部或特定基因的表观遗传具有影响。
我们不知道什么?
·大多表观遗传改变仅仅与变态反应性疾病具有相关性,目前还不清楚哪些变化是引起疾病的或者可能具有功能的相关性。
·因为表观遗传改变具有细胞特异性,哪个是应该作为研究和靶点的相关细胞型?
·个性化治疗,例如嗜酸性粒细胞清除策略临床表现出实用性,但是仍不知道仅仅是消除嗜酸性粒细胞(相应的疾病症状改善)还是逆转嗜酸性粒细胞表观遗传状态变化而恢复到健康状态。
·目前已经确定了表观遗传变化,但是仍处于初期阶段,大多数研究由于统计效力和可重复性不足而具有局限性。
·表观遗传编辑带来了很大希望,然而,细胞类型特异性、脱靶效应和耐久性在应用于临床干预之前仍存在许多挑战。
·在临床实践应用之前,需要进一步研究EVs的来源、形成机制和细胞类型特异性。
参考文献请见原文:
J Allergy Clin Immunol,2018,142: 716-726
专家点评
高金明中国医学科学院北京协和医学院北京协和医院呼吸科
表观遗传学是指在基因转录或翻译水平,非DNA变异导致了基因表达水平以及改变的表型具有“可遗传的”特点,包括DNA甲基化、组蛋白修饰和小非编码RNA调控等。变态反应和哮喘是环境因素与基因相互作用的结果,环境因素诸如感染、食物、被动吸烟和变应原可以导致基因的错误表达或功能异常。表观基因组关联研究表明,基因表达的表观遗传调节和变态反应以及哮喘具有相关性。该综述全面总结了上述表观遗传调控以及微生物组和表观遗传组之间相互作用等在变态反应性疾病的发病以及免疫调节中的作用。非过敏变态反应性疾病的临床研究显示,miRNA类似物和抑制剂、人工合成反义分子具有良好的效果。表观遗传标志物有望用于监测疾病病情、筛选适合患者个体的药物、剂量和方式。作为分子靶位,逆转异常的表型修饰可能是变态反应性疾病治疗的新途径。变态反应的表观遗传学研究仍处于早期阶段,还不清楚表观遗传学的哪些变化与疾病相关。表观遗传改变具有细胞特异性,应该以哪种细胞类型为研究靶点尚不清楚。关于个性化治疗,以嗜酸性粒细胞为例,特别需要知道是应该消除嗜酸性粒细胞还是逆转嗜酸性粒细胞表观遗传性状使之恢复到健康状态。关于表观遗传编辑,诸如细胞类型特异性、脱靶效应和耐久性在应用于临床之前仍存在许多挑战。

高金明
北京协和医学院特聘教授,协和学者,博士生导师。毕业于天津医学院医学系,在北京协和医学院获得硕士、博士学位,在哈佛医学院完成博士后训练。历任北京协和医院内科住院医师和总住院医师,呼吸内科主治医师和副主任医师,目前是呼吸科主任医师。主要研究方向:气道炎症性疾病的发病机制和干预治疗。作为第一完成人获2009年中华医学科技奖三等奖(支气管哮喘的基础和临床研究);入选“教育部新世纪优秀人才支持计划”;主持国家自然科学基金等多项科研课题,主持OM-85预防支气管扩张症急性加重临床多中心研究。以通信作者或第一作者在RedJournal、JBC、Thorax等SCI杂志发表论文多篇。
猜你喜欢
保健医苑(2022年5期)2022-06-10
中国典型病例大全(2022年10期)2022-05-10
昆明医科大学学报(2022年1期)2022-02-28
世界科学技术-中医药现代化(2021年8期)2021-12-21
河北果树(2021年4期)2021-12-02
昆明医科大学学报(2021年5期)2021-07-22
当代水产(2021年4期)2021-07-20
中学生物学(2020年10期)2020-12-25
福建基础教育研究(2019年10期)2019-05-28
医学研究杂志(2015年12期)2015-06-10
