
Novel_miR218对山羊外周血单核细胞中SLAM受体表达的影响
2018-11-30 07:14宋华杰王晶钰齐雪峰
畜牧兽医学报 2018年11期
宋华杰,王 婷,李 珍,王晶钰,齐雪峰
(西北农林科技大学动物医学院,杨凌 712100)
小反刍兽疫(Peste des petits ruminants,PPR)是由小反刍兽疫病毒(Peste des petits ruminants virus,PPRV)引起山羊、绵羊及野生小反刍兽的一种急性、烈性、接触性动物传染病。该病至今依然是世界范围内严重威胁动物健康养殖的重大疫病之一。PPRV对淋巴细胞和上皮细胞具有较强噬性,其感染宿主后造成机体免疫抑制是其致病的主要特征[1]。虽然目前对于PPRV已经做了很多有价值的研究工作,但关于该病毒对山羊致病机制研究仍不十分明了。淋巴细胞信号活化分子(signaling lymphocyte activation molecule,SLAM)表达于多种哺乳动物淋巴细胞、树突细胞及巨噬细胞等,是PPRV等麻疹病毒属病毒在淋巴细胞的主要受体[2-6]。山羊的外周血单核细胞(peripheral blood mononuclear cells,PBMCs)被广泛用于小反刍兽疫病毒致免疫抑制相关研究。研究表明,山羊PBMCs中SLAM受体表达水平与PPRV的增殖水平密切相关[4,7],而有关SLAM受体表达调节机制及其在PPRV感染与复制中的具体作用尚不完全清楚。
近年来,越来越多的证据表明宿主细胞小RNA(miRNA)是机体先天性/获得性免疫以及宿主-病原体相互作用的复杂网络中不可或缺的调节因子,其在抗病毒感染和复制方面发挥着重要作用[8-10]。Novel_miR218是本实验室在山羊PBMCs中新发现的一种微小核糖核酸分子,基于RNAhybrid及miRanda等生物信息学分析表明SLAM是novel_miR218的潜在靶基因,但关于novel_miR218对山羊PBMCs中SLAM表达水平调节机制尚需进一步证实。
本研究旨在研究novel_miR218在PPRV感染山羊PBMCs中对SLAM受体表达水平的调节作用及其对病毒增殖的影响,研究结果对进一步阐明PPRV在羊源细胞中的复制及其致病机制具有重要的理论意义和潜在的应用价值。
1 材料与方法
1.1 病毒及细胞
所用PPRV标准疫苗毒株N75-1为笔者实验室保存,病毒在Vero细胞上增殖,滴度是105.65TCID50·mL-1;山羊PBMCs从西北农林科技大学实验动物中心18月龄健康莎能奶山羊颈静脉处无菌采血,随后用淋巴细胞分离液分离获得,通过含有100单位的青霉素、100 μg·mL-1的链霉素和10%胎牛血清的RPMI 1640培养基培养。
1.2 主要试剂
DMEM/F-12为Gibco产品;胎牛血清为Hyclone产品;胰蛋白酶为Amresco产品;Lipofectamine RNAiMAX转染试剂、RNA酶抑制剂和TRIzol RNA试剂盒均为Invitrogen公司产品;逆转录试剂盒和miRNA定量试剂盒均为广州锐博生物科技有限公司产品;鼠抗SLAM单克隆抗体、兔抗β-actin抗体、HRP-羊抗兔IgG和HRP-羊抗鼠IgG均购自Invitrogen公司;鼠抗PPRV N蛋白抗体由中国兽医药品监察所提供。
1.3 引物设计与合成
参照NCBI公布的山羊SLAM基因序列设计引物,其正向引物序列:5′-AACTGGAGTGAGGAAGCAGGT-3′,反向引物序列:5′-CGCAATGCAGATGTAGACGTT-3′;内参β-actin的正向引物序列:5′-GCATTGCCCTCAACGACCACTTTGTC-3′,反向引物序列:5′-CTCCTTGGAGGCCATGTGGACCATG-3′。Novel_miR218序列信息为5′-CUCCCAGCGCUGUCACCAC-3′,其定量PCR引物序列购自广州锐博生物科技有限公司(产品编号:miR8003168、miR8003167)。
1.4 Novel_miR218 mimic及inhibitor序列
Novel_miR218模拟物(novel_miR218 mimic)、novel_miR218拮抗物(novel_miR218 inhibitor)均购自广州锐博生物科技有限公司(产品编号:miR1170901031458、miR2170901031600)。
1.5 病毒感染
山羊PBMCs分离后在5%CO2浓度的培养箱、10%胎牛血清的RPMI 1640培养基中培养16 h。未贴壁的山羊PBMCs在病毒接种前用RPMI 1640培养基轻轻洗去。之后用MOI=1的PPRV N75-1株病毒接种山羊PBMCs,同时设置未感染对照,于感染后24 h(hpi)收样。感染组和未感染对照组均设置3个独立的生物学重复。
1.6 RNA的提取和荧光定量PCR
用TRIzol reagent处理各组PBMCs后提取总RNA,Nanodrop2000超微量分光光度计定量,用cDNA逆转录试剂盒将RNA逆转录为cDNA,采用SYBR GreenⅠ法进行qRT-PCR,每组3个重复,检测PPRV感染山羊PBMCs中SLAMmRNA和novel_miR218的表达水平变化。SLAMmRNA和novel_miR218检测分别以β-actin和5S为内参基因,2-ΔΔCt法进行相对定量计算,每个样品重复3次求平均值。
1.7 Western blot检测
将作用不同时间段的各组细胞用RIPA裂解后进行SDS-PAGE,然后将蛋白质电转至PVDF膜上,5%脱脂奶粉室温封闭2 h,分别用SLAM抗体、PPRV N蛋白抗体、β-actin抗体在4 ℃孵育过夜,TBST洗膜4次,二抗孵育1 h,TBST洗膜6次,加入化学发光剂ECL,曝光获得相应条带,采用ImageJ2x 软件分析灰度值。
1.8 转染novel_miR218的模拟物和拮抗物
按照Invitrogen公司提供Lipofectamine RNAiMAX转染试剂说明书,分别将novel_miR218模拟物(novel_miR218 mimic)、novel_miR218 拮抗物(novel_miR218 inhibitor)及相应的对照miRNA转染入细胞中,置于37 ℃、5% CO2细胞培养箱中孵育48 h;之后分别于各孔接种1.0 MOI值PPRV,于37 ℃培养24 h;通过Western blot检测转染细胞中SLAM和PPRV N蛋白的表达水平。每个试验设3个重复,同时设定对照组。
1.9 数据处理与统计

2 结 果
2.1 PPRV在山羊PBMCs中的增殖规律
在PPRV感染山羊PBMCs后不同时间,通过观察细胞CPE及检测病毒N蛋白的表达量明确了PPRV N75-1毒株在山羊PBMCs中的复制规律。与对照组相比,PPRV感染细胞可见细胞肿胀、聚集及脱落等细胞病变,且随着感染时间延长细胞病变越来越明显(图1)。Western blot结果显示,PPRV N蛋白的表达水平在24 hpi时明显增加且随着感染时间延长而不断升高(图2)。PPRV感染山羊PBMCs中SLAM蛋白的表达水平检测表明,在24和48 hpi时SLAM表达水平较0 hpi时极显著增加(P<0.01),而在72和96 hpi时表达水平降低(图2)。

图1 PPRV感染山羊PBMCs后的细胞病变
Fig.1 CPE of PPRV-infected goat PBMCs

与对照组(0 hpi)比较,*.P<0.05; **.P<0.01
Compared with control (0 hpi), *.P<0.05; **.P<0.01
图2 Western blot检测PPRV感染山羊PBMCs中相关蛋白的表达
Fig.2 Western blot analysis of protein relative expression of PPRV-infected goat PBMCs
2.2 qRT-PCR检测SLAM mRNA和novel_miR218表达水平
PPRV感染山羊PBMCs中SLAMmRNA和novel_miR218的qRT-PCR检测结果表明, 24 hpi时SLAMmRNA表达水平较0 hpi极显著增加(P<0.01),随后逐渐降低;对照组SLAMmRNA表达水平在各检测时间点均较低(图3)。Novel_miR218表达水平变化趋势则与SLAMmRNA呈负相关,24 hpi时novel_miR218表达水平较0 hpi极显著降低(P<0.01),随后逐渐升高;在24~96 hpi,PPRV感染组novel_miR218表达水平均显著低于同时间点对照组(图4)。
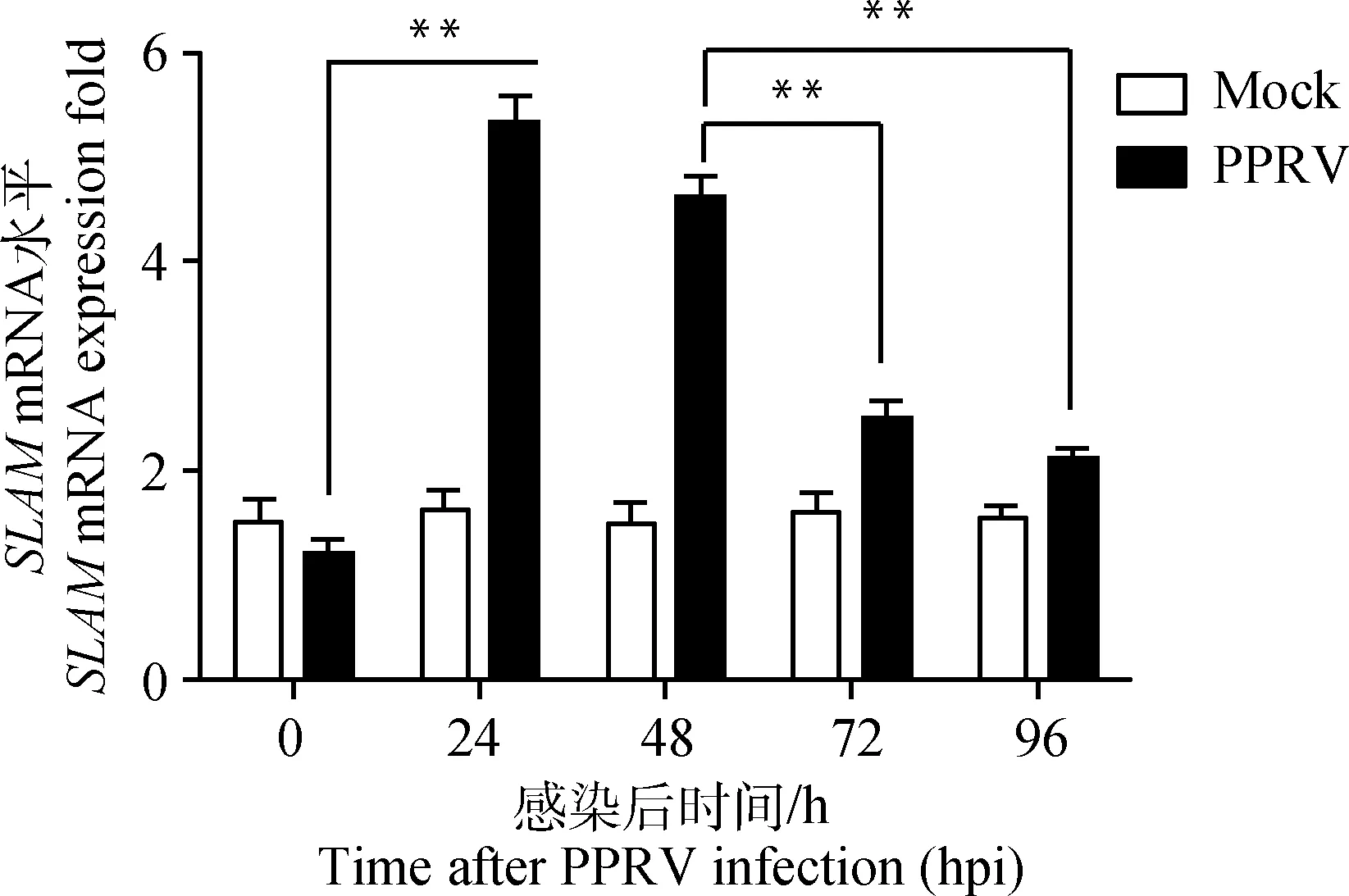
与对照组(0 hpi)比较,*.P<0.05; **.P<0.01
Compared with control (0 hpi), *.P<0.05; **.P<0.01
图3 qRT-PCR检测PPRV感染山羊PBMCs后SLAM mRNA水平
Fig.3 qRT-PCR analysis of SLAM mRNA expression fold of PPRV-infected goat PBMCs

与对照组(0 hpi)比较,*.P<0.05; **.P<0.01
Compared with control(0 hpi), *.P<0.05; **.P<0.01
图4 qRT-PCR检测PPRV感染山羊PBMCs后novel_miR218水平
Fig.4 qRT-PCR analysis of novel_miR218 expression fold of PPRV-infected goat PBMCs
2.3 转染novel_miR218模拟物和拮抗物对SLAM和PPRV N蛋白的影响
如图5所示,novel_miR218 mimic转染组中SLAM和PPRV-N蛋白表达水平均较转染对照组降低(P<0.05 或P<0.01),且呈转染浓度依赖性;相反地(图6),novel_miR218 inhibitor转染组中SLAM和PPRV-N蛋白表达水平均较对照组呈转染浓度依赖性升高(P<0.05或P<0.01)。表明novel_miR218对SLAM受体的蛋白表达具有明显负调控作用,同时影响PPRV的增殖水平。
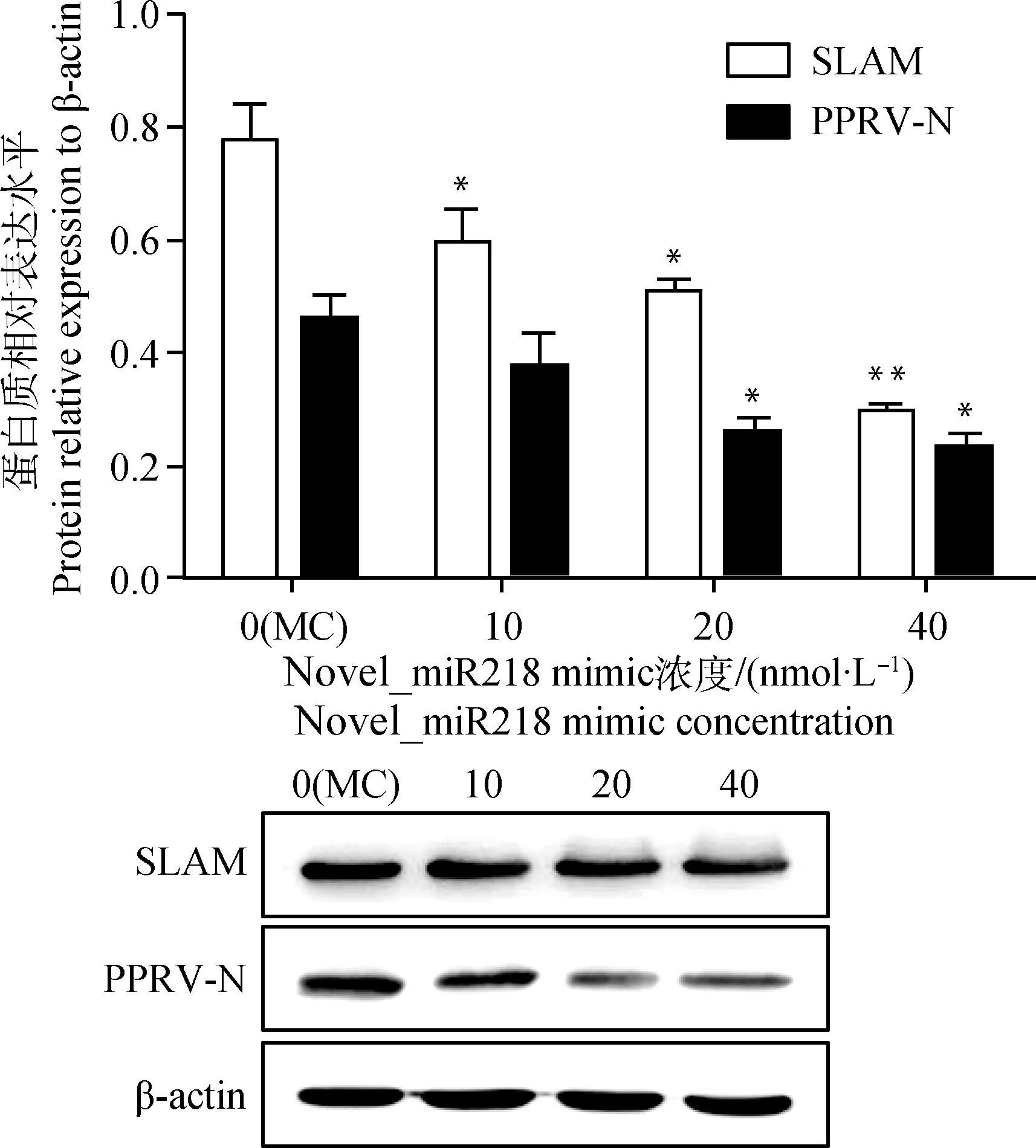
与对照组(MC)比较,*.P<0.05;**.P<0.01
Compared with control(MC), *.P<0.05; **.P<0.01
图5 Western blot检测转染novel_miR218 mimic后PPRV感染山羊PBMCs中相关蛋白表达水平
Fig.5 Western blot analysis of protein relative expression after the transfection of novel_miR218 mimic into PPRV-infected goat PBMCs

与对照组(IC)比较,*.P<0.05; **.P<0.01
Compared with control(IC), *.P<0.05; **.P<0.01
图6 Western blot检测转染novel_miR218 inhibitor后PPRV感染山羊PBMCs中相关蛋白表达水平
Fig.6 Western blot analysis of protein relative expression after the transfection of novel_miR218 inhibitor into PPRV-infected goat PBMCs
3 讨 论
小反刍兽疫是由副黏病毒科麻疹病毒属小反刍兽疫病毒(PPRV)引起的一种急性、热性、高度接触性一类传染病,主要感染包括野生动物在内的小反刍动物,其中山羊高度易感,绵羊易感。该病危害严重,其发病率可高达90%~100%,病死率可达50%~100%[11-13]。 与其他病毒一样,PPRV侵染宿主细胞的“核心”机制是病毒与特定受体结合进而感染宿主细胞[14-15]。然而,PPRV对感染宿主的致病机制尚不十分明了,特别是在山羊等高度易感动物。现已确定PPRV的天然受体有两个:SLAM和脊髓灰质炎病毒受体(Nectin4/PVRL4)。其中SLAM仅仅在T淋巴细胞、巨噬细胞、NK细胞和树突状细胞等免疫细胞表面表达,Nectin4则主要在上皮细胞中表达[16-18]。SLAM是PPRV感染宿主并致病的关键受体,当PPRV通过呼吸道途径侵染宿主时,它首先与巨噬细胞和树突状细胞表面SLAM受体发生结合并相互作用,而后被转运到局部淋巴结进行增殖扩散[19-23]。PPRV的N结构蛋白被广泛用于对PPRV增殖规律的研究[15,23-24]。2008年,Pawar等[4,24]在通过siRNA干扰技术证实SLAM是PPRV感染的病毒受体的基础上,进一步研究发现山羊外周血单核细胞中SLAM的表达水平与PPRV复制具有很高的相关性,不同表达水平的SLAMmRNA影响病毒的复制和滴度。然而,目前对于SLAM表达水平的调控机制尚不清楚。
越来越多的研究表明[25-26],miRNAs与病毒的感染和致病有密切联系,发挥着重要功能。在麻疹病毒属的研究表明[27-28],麻疹病毒感染可导致宿主细胞中miRNA表达谱及其相应的靶基因mRNA表达发生显著变化。此外,Geekiyanage和Galanis[29]研究表明,miR-31和miR-128可通过调控麻疹病毒在肿瘤细胞的受体Nectin4,进而调控该病毒对该细胞的易感性。本实验室在对PPRV感染山羊PBMCs中miRNA表达谱进行差异分析表明,novel_miR218有可能对SLAM受体的表达具有一定调节作用。本研究中通过检测PPRV感染山羊PBMCs不同时间后SLAM和novel_miR218表达水平发现,novel_miR218表达水平变化趋势与SLAMmRNA呈负相关,而转染novel_miR218 mimic后SLAM和PPRV-N蛋白表达水平均较转染对照组降低,转染novel_miR218 inhibitor后SLAM和PPRV-N蛋白表达水平均较转染对照组升高,以上结果表明novel_miR218对SLAM受体的蛋白表达具有明显负调控作用。因此PPRV在早期侵染过程中,可能通过novel_miR218介导对SLAM受体表达丰度的调控,进而影响PPRV在山羊PBMCs中的增殖水平。另外,本研究结果是否与PBMCs分离山羊种属、性别等有关有待进一步研究。现已证实,除了麻疹病毒属病毒外,其他多种病毒及致病菌也均以SLAM蛋白作为其在宿主细胞的感染受体并通过调控该受体表达水平进而达到免疫逃逸[30]。因此,关于SLAM受体表达及其相关信号通路调控研究对深入了解相关病原体致病机制具有重要意义。
4 结 论
PPRV感染山羊PBMCs中SLAM受体表达水平与novel_miR218的表达水平呈负相关,其中SLAM受体表达呈先升高后降低趋势,而novel_miR218的表达呈先降低后升高趋势;山羊PBMCs源novel_miR218对SLAM受体表达具有负调控作用并对PPRV增殖水平有一定影响。本项目研究结果对进一步阐明PPRV在羊源细胞中的复制及其致病机制等均具有重要的理论意义和潜在的应用价值。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
今日农业(2021年15期)2021-11-26
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
江西医药(2018年8期)2018-10-24
作文周刊·小学一年级版(2017年46期)2018-01-16
小天使·一年级语数英综合(2017年4期)2017-04-18
小星星·阅读100分(高年级)(2015年11期)2015-11-28
医学研究杂志(2015年12期)2015-06-10
