
桉叶油含药血清联合维甲酸对体外培养神经干细胞Pax3及Cx43的影响*
2018-11-15 02:21:30胡晓芳余永莉张彬渝刘茂生苏丽莉遵义医学院珠海校区人体解剖与组织胚胎学教研室广东珠海519041
现代医药卫生 2018年21期
胡晓芳,余永莉,陈 涛,张彬渝,刘茂生,苏丽莉(遵义医学院珠海校区人体解剖与组织胚胎学教研室,广东珠海519041)
桉叶油是从桉树叶中提取的具有芳香味的挥发性油,具有抗菌、抗氧化、抗炎、促渗透、杀虫、驱蚊、止痒及防腐作用[1]。有文献报道,桉叶油能舒张气管和血管平滑肌、心肌,其作用机制可能是影响膜上离子通道的离子流[2-4]。本课题组前期研究发现,桉叶油对维甲酸(RA)导致胎鼠神经管缺陷有一定的拮抗作用,其机制可能与桉叶油拮抗RA上调胎鼠神经组织的Pax3、Cx43蛋白过度表达有关。本研究利用桉叶油制备含药血清,联合RA作用体外培养神经干细胞,通过检测细胞Pax3、Cx43基因表达水平,从体外实验探讨桉叶油是否也具有拮抗RA上调胎鼠神经组织的Pax3、Cx43蛋白表达的作用。
1 材料与方法
1.1 材料
1.1.1 实验动物 选择SPF级SD大鼠,体重180~220 g[购自广东省医学实验动物中心,实验动物合格证号编号:0094460,许可证号:SCXK(粤)2008-0002];饲养条件:室温24~27℃,通风,相对湿度68%~80%,每天光照12 h,自由摄食、饮水。
1.1.2 药品及试剂 RA粉末原料药(北京贝利莱斯生物化学有限公司,批号:110801),桉叶油[桉叶素99.5%,德信行(珠海)香精香料有限公司,批号:110626];选用金龙鱼花生油作为药品溶剂。
1.2 方法
1.2.1 桉叶油含药血清的制备 选择体重250~300 g清洁级的雄性SD大鼠,将其随机分为6组,每组3只,分别作为正常血清组(大鼠自由饮食)、溶剂血清对照组(每只大鼠早晨灌胃花生油3 mL,每天1次,连续灌胃 7 d)、桉叶油血清组(分别用桉叶油 300、1 000、1 500 mg∕kg 于早晨灌胃,每天 1 次,连续灌胃 7 d)。各组末次灌胃后2.5 h左右,用10%的水合氯醛进行麻醉,解剖胸腔,用一次性无菌注射器心脏采血,离心,收集血清。无菌滤器过滤除菌,置于56℃水浴30 min灭活,-20℃保存备用。
1.2.2 神经干细胞培养及鉴定 在无菌条件下取24 h内新生SD大鼠的海马组织,轻微机械吹打组织,用200目过滤网过滤、收集单细胞悬液。加入适量DMEM∕F12 全培养液[内含 2% 维生素 B27、20 μg∕L 成纤维细胞生长因子(FGF)和 20 μg∕L 表皮生长因子(EGF)],放入37℃、5%CO2饱和湿度CO2培养箱中换液培养7 d,传代1次,收集第2代神经干细胞,轻微吹打、吹散细胞球,制成单细胞悬液,调整细胞密度为1×105mL-1接种于6孔培养板,每孔4 mL,分为BrdU标记组和对照组。BrdU 标记组加入 BrdU 10 μmol∕L,对照组加入 DMEM∕F12全培养液,培养48 h收集各孔细胞,用4%的多聚甲醛固定细胞后,采用免疫荧光法检测BrdU、Nestin。鉴定BrdU组一抗为兔抗鼠BrdU抗体,鉴定Nestin组一抗为兔抗鼠Nestin抗体,两组二抗均为羊抗兔异硫氰酸荧光素-免疫球蛋白G(FITC-IgG)。并设立一抗为磷酸盐缓冲溶液(PBS)的阴性对照组。使用荧光显微镜观察结果。
1.2.3 桉叶油含药血清对体外培养胎鼠神经干细胞Pax3、Cx43表达的影响 收集第二代神经球细胞,加入新的无血清DMEM∕F12全培养液,轻微吹打、吹散细胞球,制成单细胞悬液,调整细胞密度为1×105mL-1接种于培养板,分为5组,分别加入自由饮食SD大鼠血清、溶剂花生油灌胃血清、桉叶油 1 500、1 000、300 mg∕kg灌胃鼠血清,各组血清终浓度为5%。各组细胞放入37℃、5%CO2、饱和湿度条件下CO2培养箱中培养48 h,收集细胞,采用蛋白免疫印迹法(Western blotting)检测细胞Pax3、Cx43蛋白表达水平,并采用逆转录-聚合酶链反应(RT-PCR)检测细胞Pax3、Cx43基因表达水平。
1.2.4 桉叶油含药血清联合RA对体外培养胎鼠神经干细胞Pax3、Cx43表达的影响 收集第2代神经球细胞,加入新的无血清DMEM∕F12全培养液,轻微吹打、吹散细胞球,制成单细胞悬液,调整细胞密度为1×105mL-1接种于6孔培养板,分为6组,正常SD大鼠血清5%+RA(5 μmol∕L),溶剂花生油含药血清 5%+RA(5 μmol∕L),桉叶油 1 500 mg∕kg 含药血清 5%+RA(5 μmol∕L),1 000 mg∕kg 含药血清 5%+RA(5 μmol∕L)、300 mg∕kg 含药血清 5%+RA(5 μmol∕L),RA(5μmol∕L)。各组细胞放入37℃、5%CO2饱和湿度条件下CO2培养箱中培养48 h,收集细胞,采用Western blotting检测细胞Pax3、Cx43蛋白表达水平,并采用RT-PCR检测细胞Pax3、Cx43基因表达水平。
1.3 统计学处理 应用SPSS13.0统计软件进行数据分析,计量资料以表示,各组均数比较采用方差分析。检测水准α=0.05,P<0.05为差异有统计学意义。
2 结 果
2.1 大鼠神经干细胞的培养及鉴定 收集新生SD大鼠海马组织细胞在无血清的DMEM∕F12全培养液培养24 h后,肉眼可见培养瓶中有乳白色小块状或者颗粒状悬浮物,镜下观察悬浮物为聚集的细胞块,细胞轮廓清晰,有光泽,其周围有悬浮的死细胞碎片,瓶底也有大量有光泽的单个细胞。换液后吹散悬浮小块继续培养,如此换液吹散培养到第7天,可见大量大小不等的神经细胞球。收集第2代细胞免疫荧光鉴定结果显示Nestin、Brdu表达呈阳性(图1),整个培养过程中无细胞贴壁现象出现。
2.2 桉叶油含药血清对体外培养胎鼠神经干细胞Pax3、Cx43蛋白及mRNA表达的影响 用5%的血清作用细胞后,桉叶油含药血清各组细胞的Cx43蛋白及基因表达水平较正常血清组的表达水平均高(P<0.05),Pax3蛋白及基因表达水平较正常血清组的表达水平均低(P<0.05)。见图2、表1。
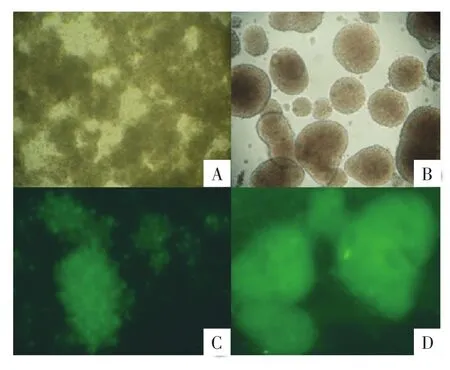
图1 神经干细胞的培养与鉴定(200×)
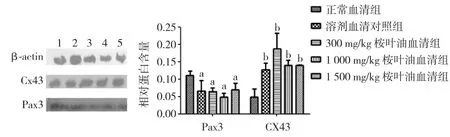
图2 桉叶油含药血清作用神经干细胞48 h Pax3、Cx43蛋白表达水平
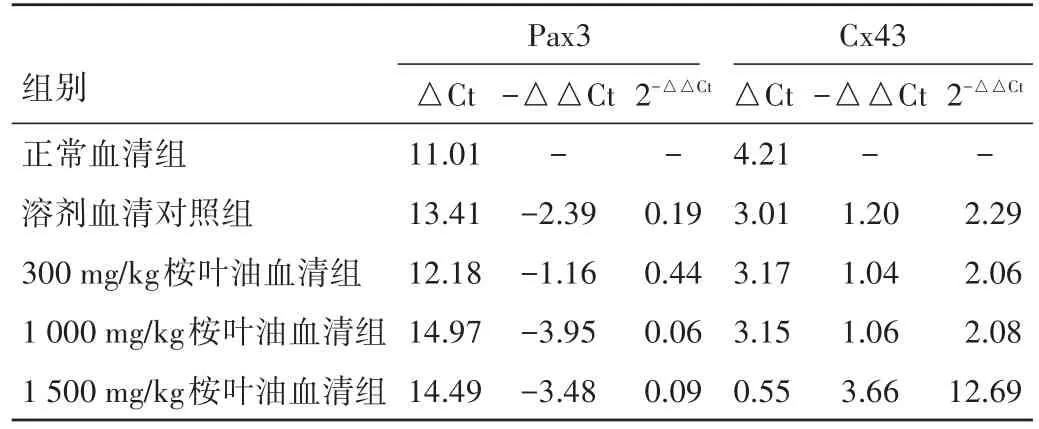
表1 桉叶油含药血清作用神经干细胞48 h Pax3、Cx43基因表达水平相对正常血清组Pax3、Cx43基因表达改变情况
2.3 桉叶油含药血清联合维甲酸对体外培养胎鼠神经干细胞Pax3、Cx43蛋白及mRNA表达的影响 用5%的血清+RA作用细胞后,与单纯RA组比较,血清各组Pax3蛋白及mRNA表达水平均降低(P<0.05);桉叶油含药血清各组的Pax3蛋白及mRNA较正常血清组表达水平均有差异(P<0.05)。
用5%的血清+RA作用细胞后,与单纯RA组比较,血清各组Cx43蛋白及mRNA表达水平均降低;但在5%的血清+RA各组中,桉叶油各组Cx43蛋白及其mRNA与正常血清组比较无显著差异(P>0.05)。结合单纯桉叶油血清可上调细胞的Cx43蛋白及其mRNA表达,但与RA联合后,其对神经干细胞Cx43的影响与正常血清组比较无显著差异(P>0.05)。见图3、表2。
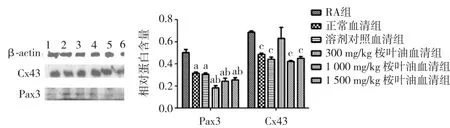
图3 桉叶油含药血清联合维甲酸作用神经干细胞48 h Pax3、Cx43蛋白表达水平
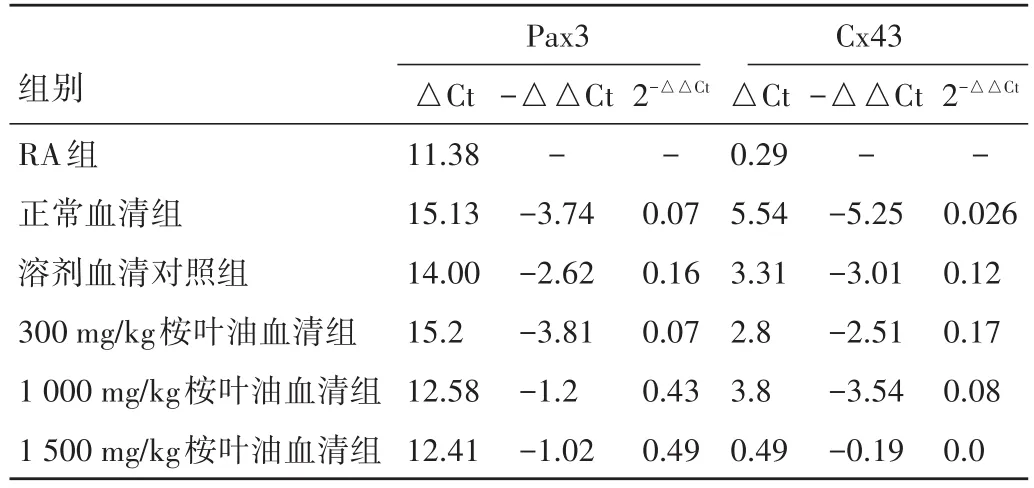
表2 桉叶油含药血清联合RA作用体外培养胎鼠神经干细胞48 h Pax3、Cx43 mRNA相对RA组Pax3、Cx43 mRNA表达水平改变情况
3 讨 论
RA是维生素A在体内氧化代谢的产物,具有多种生物学功能,在胚胎生长发育过程中对胚胎的形成、细胞生长与分化起重要的调节作用。RA是一种很强的诱导分化剂,可诱导300多种功能蛋白质的表达。已有文献报道,小剂量RA可诱导小鼠胚胎干细胞分化为神经元。钟德君等[5]观察不同浓度的RA在鼠胚神经干细胞增殖和分化中的作用,发现RA具有显著地促进大鼠神经干细胞(NSCs)分化为神经元的作用,且 1.0 μmol∕L RA 诱导分化效果最佳。蒋震伟等[6]用 10 μmol∕L RA 处理人胚神经干细胞后,将神经干细胞移植入受损的大鼠脊髓内,发现神经干细胞在体内能替代缺失的神经元,能促进受损的脊髓功能恢复。然而,RA也是致畸因子,过量的RA可导致实验动物的神经管畸形、腭裂、骨质疏松等等多种畸形。蔡炜嵩等[7]利用RA灌胃造大鼠神经管畸形动物模型观察其出现神经组织畸胎瘤、脊髓裂开、脊柱裂等畸形形态学特点,发现RA可干扰神经发育导致脊柱裂、脊髓裂开和神经错构瘤的发生。李红丽等[8]用RA建立小鼠神经管畸形模型,体外培养神经干细胞并检测其增殖变化,发现胚胎早期神经干细胞的数目减少及神经细胞增殖受抑制与RA致神经管畸形的发生有密切关系。由于胚胎畸形发生是先天性的,一旦发生后具有不可逆性,故其治疗方法主要以预防性治疗为主。不少学者利用RA造胚胎致畸模型,通过研究药物对其的拮抗作用,来寻找有效预防治疗先天畸形的方法。本课题组前期研究中利用桉叶油联合RA灌胃SD孕鼠,探讨其在预防性治疗胚胎畸形中是否具有药用价值,结果发现,桉叶油对RA引起胎鼠畸形,生长发育迟缓,骨骼发育及骨化迟缓有一定的拮抗作用[9-10],对胎鼠脑组织的Cx43、Pax3蛋白和基因表达水平进行检测,发现RA组胎鼠神经组织的Pax3、Cx43蛋白过度表达,桉叶油具有拮抗RA上调胎鼠神经组织的Pax3、Cx43蛋白表达的作用,但其具体机制尚不清楚。
Cx43是缝隙连接蛋白家族的重要成员,也是脊椎动物中表达最多、最广泛的缝隙连接蛋白。缝隙连接又称通讯连接,是细胞间物质交换和信息传递的通道。该通道允许离子、小分子物质等自由通过,传递细胞间的信息和能量,对细胞的新陈代谢、内环境稳定、细胞增殖、分化、生长调控等各方面起着重要作用。在神经系统和循环系统尚未建立或未发育完善的早期胚胎,长距离的信息传递难以进行,胚胎各部分细胞的增殖、分化与生长发育离不开细胞间离子信息的交换,缝隙连接异常将导致细胞分化迁移异常。有研究表明,Cx43表达及其缝隙连接功能的异常与许多神经系统疾病的病理生理过程有密切的联系[11]。REAUME等[12]发现,Cx43无义突变可造成信息传递缺陷,影响神经细胞的迁移、增生及细胞凋亡过程。喻博等[13]用体外培养的胎鼠神经干细胞移植治疗大鼠脑损伤,发现神经干细胞移植对脑损伤的恢复有一定作用,且与CX43的表达水平有关。WATANABE等[14]研究认为,过量RA是通过增加Cx43 mRNA的表达上调Cx43蛋白的表达,使信息传递发生紊乱,从而阻止细胞生长。本研究用5 μmol∕L RA作用神经干细胞48 h,也发现RA具有上调细胞Cx43 mRNA及蛋白表达的作用,5%桉叶油含药血清联合RA作用细胞48 h后,细胞的Cx43 mRNA及蛋白表达与单纯RA组比较均显著降低,提示桉叶油含药血清具有拮抗RA上调细胞Cx43 mRNA及其蛋白表达的作用,而单纯的5%桉叶油含药血清作用神经干细胞48 h后,细胞的Cx43 mRNA及其蛋白表达水平增加,此结果进一步证明桉叶油含药血清对RA上调细胞Cx43表达有拮抗作用,而非协同作用。Pax3基因属于Pax基因家族,是重要的转录调控因子,在胚胎发育过程中对组织和器官的分化起着重要的调控作用。STOYKOVA等[15]研究发现,在神经管细胞开始分化之前,在整个神经管轴的顶板、侧翼板及神经嵴细胞均有表达Pax3基因已表达。Pax3表达过高或过低均可导致相关疾病的发生,WU等[16]研究证明,神经嵴细胞中Pax3的持续高表达会导致小鼠腭裂和骨畸形。ZHOU等[17]研究发现,Pax3能调节Soostdc1的表达从而阻止神经嵴对骨形成蛋白的诱导,导致胚胎出现脊柱裂或脑膨出。BURREN等[18]研究发现,叶酸缺乏可致小鼠Pax3功能丧失,从而导致神经管畸形。王涛等[19]用RA及牛磺酸建立小鼠神经管畸形及干预模型,发现RA的致畸作用与Pax3基因表达的时相差异性有关。KENNEDY等[20]用低剂量的RA处理P19或小鼠胚胎干细胞,发现RA是通过上调Wnt3a、PAX3、Meox1、MyoD 和 Myogenin的表达,实现骨骼肌的成肌增强。本研究用5 μmol∕L RA作用神经干细胞48 h后,细胞的Pax3的表达也是增加的,RA联合5%的桉叶油含药血清作用细胞48 h后,细胞的Pax3蛋白及mRNA表达水平均降低,提示桉叶油含药血清具有拮抗RA上调细胞Pax3蛋白和mRNA表达的作用。
本研究表明,RA具有上调神经干细胞Cx43、Pax3的表达,桉叶油含药血清具有拮抗RA上调神经干细胞Cx43、Pax3表达的作用。此结论与本课题组前期体内实验研究结论一致,从而进一步证明了影响Cx43、Pax3的表达可能是桉叶油含药血清具有拮抗RA致畸作用的机制之一。
猜你喜欢
电镀与涂饰(2023年14期)2023-08-08 08:12:04
家庭医学(下半月)(2020年4期)2020-05-30 12:42:52
中成药(2018年10期)2018-10-26 03:40:44
天然产物研究与开发(2016年11期)2016-06-15 20:29:17
中国实验诊断学(2016年1期)2016-02-29 03:10:37
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
山西医科大学学报(2014年1期)2014-11-20 09:45:52
癌变·畸变·突变(2014年1期)2014-03-01 04:39:35
山东医药(2013年7期)2013-06-14 06:36:56
食品科学(2013年8期)2013-03-11 18:21:13
