
紫耳箭竹克隆形态可塑性对典型冠层结构及光环境的响应
2018-11-14 12:46:20黄慧敏向运蓉何丹妮陶建平
生态学报 2018年19期
黄慧敏,董 蓉,钱 凤,向运蓉,何丹妮,陈 淼,陶建平
西南大学生命科学学院,三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室, 重庆 400715
形态可塑性是同一基因型的植物个体通过改变自身的形态而对环境变化做出相应变化的能力,是生物适应环境的一种方式,即形态可塑性是环境对基因型表达的一种修饰[1]。由于自然环境中异质性是普遍存在,种群的维持与更新与其形态可塑性紧密相关[2]。同时,在克隆植物中形态可塑性也是其适应异质性环境的重要途径。在不同资源水平下,克隆植物可以通过形态塑造来适应不同的资源水平[3- 5],主要表现为构件、资源吸收结构形态特征与生物量分配等变化[6- 7]。近些年有关克隆植物个体行为对环境的响应及其在生态系统中所起到的作用越来越受到人们的关注[8],而其形态可塑性的适应性响应与物种入侵能力、物种进化和物种共存等方面又有着重要的关系[9- 10]。
小径竹类是多年生一次性开花结实的木本克隆植物,广泛分布于亚高山暗针叶林及亚热带山地阔叶林等异质性强的生境中,为森林灌木层的优势种甚至单优种[11]。已有研究表明,小径竹在光照强度相差甚远的林外、林缘、林窗和林下等生境下均有分布[6,12]。不同的林冠环境对小径竹的生长具有一定的影响,尤其是光环境[13- 16]。小径竹可以通过改变分株数目、形态、生物量分配以及聚集强度和空间占据能力上,对林冠环境的差异作出有效的响应[17- 19]。在林冠郁闭度大的环境中,小径竹分株表现出比叶面积、叶面积率,叶生物量分配率增大的特征,有效适应弱光环境[20- 21]。但这些研究多集中在林隙、林窗和林缘等特殊冠层对小径竹生长的影响[12,22- 23],关于典型群落类型的冠层结构(简称典型冠层结构)对林下小径竹生长影响的研究较少,也并未量化典型群落冠层结构的特征及其所造成的光环境异质性对小径竹生长的影响。
森林冠层结构由复杂的叶、枝和树干在空间排列组成,是森林与外界环境相互作用最直接和最活跃的界面层[24]。其中,冠层结构与林下光照时空分布的关系一直是森林生态学家研究的热点。当林冠层的结构发生改变时,林下光照条件会发生不同程度的改变[25- 26]。而光照又是植物能否生存和生长的关键,因此冠层结构与林下草本层(包括更新幼苗)和灌木层的物种丰富度、物种多样性有着密切的关系[27]。目前关于群落冠层结构与林下植物的研究也多集中于此[28- 29],但在典型的群落冠层结构下,单一优势物种的生长状况如何?冠层结构及光环境发生改变时,植物如何适应性获取资源从而扩展生存?冠层结构与其适应性特征的关系又如何?这些问题目前尚未得到解答。
因此,本文拟通过对3种典型群落类型下广泛生长的克隆植物——紫耳箭竹(Fargesiadecurvata)为研究对象,比较不同冠层结构及光环境下,紫耳箭竹种群的形态特征、生物量积累和生物量分配特征的差异;探讨群落的内部是否可以通过调控冠层结构及其光环境的改变,影响小径竹的适应性能力——形态可塑性,从而可以达到协调和控制小径竹种群的发展。
1 研究地概况
金佛山国家自然保护区(28°46′—29°38′N,106°54′—107°27′E),位于重庆市南川区,属亚热带温润季风气候,云雾多,日照少,雨量充沛,湿度大。山体主要由灰岩和石灰岩组成,属于典型的喀斯特地貌。山体上部多年平均气温8.5℃,年平均降雨量约为1434.5 mm;山体下部平均气温16.6℃,年平均降雨量约1286.5 mm。海拔约为1400—2252 m,高程差大,植被类型丰富,主要森林类型为山地阔叶林(落叶阔叶林、常绿落叶阔叶混交林、常绿阔叶林)。在本研究区域内,落叶阔叶林树种主要有雷公鹅耳枥(Carpinusviminea)、糙皮桦(Betulautilis)、石灰花楸(Sorbusfolgneri)、青榨槭(Acerdavidii)等,常绿阔叶林树种主要有粗脉杜鹃(Rhododendroncoeloneurum)、金山杜鹃(Rhododendronlongipesvar.chienianum)、润楠(Machiluspingii)等,而这些林下均广泛分布有紫耳箭竹[30],为灌木层优势种。
2 材料与方法
2.1 研究材料
紫耳箭竹,箭竹属,短三角鞘系,多年生一次性开花结实的木本克隆植物[31]。地下茎属复轴型,以游击型或密集型的方式进行繁殖扩展。主要生长在海拔1150—1610 m的地方,在西南地区山地阔叶林内广泛分布,是大熊猫主要采食的竹种之一[16]。
2.2 研究方法
研究样地位于金佛山国家自然保护区的烂坝菁(29°01′N,107°09′ E)。2016年11月,选择立地条件基本一致的3种典型群落类型:落叶阔叶林、常绿落叶阔叶混交林(以下简称混交林)和常绿阔叶林(依次表示为De,Ev+De,Ev)作为研究样地。每个林型面积不小于1 hm2,间隔不小于50 m。每个林型内分别设置20 m×20 m大样方3个,共计9个大样方。对于同一种林型3个重复样方的选择,在主要物种组成、林冠郁闭度及海拔等特征尽可能保持一致。样地基本特征见表1。
2.2.1 紫耳箭竹分株种群指标测量
在各林型的每个大样方内随机设置3个5 m×5 m小样方,共计27个小样方,分别调查小样方内紫耳箭竹的分株数目、高度和盖度。随后,在每种林型下随机选择10—15个紫耳箭竹竹丛,挖取地下部分的竹鞭(由于条件的限制,无法挖取完整的基株),用水冲洗干净,记录每个竹丛中分株数目、地下茎分枝数、分枝角度、隔离者数量和笋芽数。然后从每个林型里所挖出的竹丛中随机选择一定数量的完整分株(落叶阔叶林、混交林、常绿阔叶林分别有126个、224个、136个分株),分为地上和地下部分进行测量。(1)地上部分:记录分株的株高、基径、秆节间数、秆节间长度、分枝数和叶片数,并分别采集每个分株上的所有叶片,压平带回实验室,测量叶面积(WinRHIZO Pro v.2004c Root Analysis System,Canada);(2)地下部分:测量隔离者长度、隔离者直径、隔离者节间数、隔离者节间长度、根长度、根粗细和根数量等。最后把每个分株分成秆、枝、叶、隔离者和根这5部分,用剪刀剪下,分别装入信封内,编号,放入60℃的烘箱72 h烘干至恒重,电子天平称量干重。
2.2.2 冠层结构和光环境指标测量
3个林型内,选择每个样方内紫耳箭竹种群调查的5 m×5 m小样方的中心作为冠层结构拍摄位点,共27个拍摄位点。使用Hemiview数字植物冠层分析系统(Canon EOS 50D相机,Sigma EX DC 4.5 mm鱼眼镜头)获取冠层结构和光环境数据。拍摄选在晴朗或阴天的天气进行。拍摄前需要用三脚架对其进行固定,拍摄高度设为1.5 m,保持镜头水平,指南针指向正北方向,每个位点拍摄3—5张照片。拍摄完毕后,每个位点选取3张,共81张照片,以JPG格式存储在内存卡中,用于后期分析。
2.3 数据处理
2.3.1 冠层结构和光环境特征
使用Hemiview仪器自带的分析系统对获得的照片进行分析,主要获得的参数包括:冠层开度(Canopy openness,CO)、叶面积指数(Leaf area index,LAI)、平均叶倾角(Mean leaf angle,MLA)、林下直射光(Direct solar radiation under canopy,Dir)、散射光(Diffuse solar radiation under canopy,Dif)和总光照(Total solar radiation under canopy,Tot)等。

表1 样地基本特征
De:落叶阔叶林,Deciduous broad-leaved forest;Ev+De:常绿落叶阔叶混交林,Evergreen and deciduous broad-leaved mixed forest;Ev:常绿阔叶林,Evergreen broad-leaved forest;SE:东南,South east;SW:西南,South west
2.3.2 紫耳箭竹分株种群性状特征
(1)形态性状:株高(Culm height,CH)、秆基径(Culm diameter,CD)、秆节间数(Internode number,IN)、秆节间长度(Internode length,IL)、比茎长(Specific culm length,SCL)=茎长/茎生物量、分枝数(Number of branch,NB)、单叶面积(Single leaf area,ILA)、分株叶片数(Leaf number per ramet,LNR)、叶面积率(Leaf area ratio,LAR)=叶面积/总生物量、比叶面积(Specific leaf area,SLA)=叶面积/叶生物量、根长度(Root length,RL)、根粗细(Root diameter,RD)、根数量(Root number,RN)、隔离者长度(Spacer length,SL)、隔离者基径(Spacer diamater,SD)、隔离者节间数(Spacer internode number,SIN)、隔离者节间长度(Spacer internode length,SIL)、比隔长(Specific spacer length,SSL)=隔离者长度/隔离者生物量、分枝角度(Branch angle of spacer,BA)、分枝强度(Branch intensity of spacer,BI)=母隔离者产生的子隔离者数量。
(2)生物量积累性状:秆生物量(Culm biomass,CM)、枝生物量(Branch biomass,BM)、叶生物量( Leaf biomass,LM)、隔离者生物量(Spacer biomass,SM)、根生物量(Root biomass,RM)、地上生物量(Above-ground biomass,AM))、 地下生物量(Under-ground biomass,UM)、总生物量(Total ramet biomass,TM)。
(3)生物量分配性状:茎秆生物量百分率(Allocation percentage to culm,CP);枝生物量百分率(Allocation percentage to branch,BP);叶片生物量百分率(Allocation percentage to leaf,LP);地下茎生物量百分率(Allocation percentage to spacer,SP);根生物量百分率(Allocation percentage to root,RP)、地上生物量百分率(Allocation percentage to above-ground,AP)、地下生物量百分率(Allocation percentage to under-ground,UP)。
各个性状特征的可塑性指数(Plasticity index)参考 Valladares等[32]的方法计算求得:某指标的可塑性指数=(最大值-最小值)/最大值。
用one-way ANOVA(P< 0.05)分析不同群落类型冠层结构和光环境特征的差异,比较不同冠层结构下紫耳箭竹分株种群的形态特征、生物量积累特征以及生物量分配特征间的差异性,并作LSD多重比较;利用Spearman相关法,分析冠层结构特征和光环境的关系;并用相关回归法对紫耳箭竹性状特征与冠层结构和光环境的关系进行分析,所有数据分析均用SPSS 22.0完成,并用Origin 9.0软件作图。
3 结果与分析
3.1 不同林型冠层结构和光环境特征
不同林型间的冠层结构和光环境特征存在差异(图1)。冠层开度、林下直射光、散射光和总光照的趋势相同,均表现为:落叶阔叶林>混交林>常绿阔叶林,而叶面积指数则表现出与它们相反的趋势,且差异显著(P<0.05)。平均叶倾角在落叶阔叶林中最大,而其他两个林型间无显著差异。Spearman相关分析(表2)中也发现,冠层开度、平均叶倾角、林下直射光、散射光和总光照,两两间均呈现为显著正相关,但它们均与叶面积指数呈现显著的负相关(P<0.05)。

图1 不同林型冠层结构和光环境特征比较(平均值±SE)Fig.1 Comparison of canopy structure and light conditions in different forest types (Means ± SE)Dif:林下散射光,Diffuse solar radiation under canopy;Dir:林下直射光,Direct solar radiation under canopy;图中不同字母表示差异显著

参数ParameterCOLAIMLADifDirTotCO1LAI-0.983**1MLA0.433*-0.520*1Dif0.984**-0.990**0.496*1Dir0.942**-0.939**0.571**0.947**1Tot0.955**-0.952**0.538*0.961**0.994**1
CO:冠层开度,Canopy openness;LAI:叶面积指数,Leaf area index;MLA:平均叶倾角,Mean leaf angle;Tot:林下总光照,Total solar radiation under canopy;*P<0.05;**P<0.01
3.2 紫耳箭竹分株种群的形态特征
不同林型下,紫耳箭竹分株种群的地上和地下形态特征均表现出显著差异(P<0.05)(表3)。地上形态中,紫耳箭竹的株高、基径、秆节间数、秆节间长度、分枝数、单叶面积、单叶生物量、分株叶片数在常绿阔叶林中均为最小,但比茎长、叶面积率、比叶面积却达到最大(P<0.05)。其中,基径、秆节间数、分枝数在落叶阔叶林中最大,而株高、秆节间长度、单叶面积在混交林中达最大(P<0.05)。地下形态中,紫耳箭竹分株种群的隔离者长度、隔离者直径、隔离者节间数、隔离者节间长度、根粗细和根数量均随着落叶阔叶林混交林常绿阔叶林依次显著减小(P<0.05)。比隔长和分枝角度在常绿阔叶林中达到最大,但分枝强度、根长度却是最小。
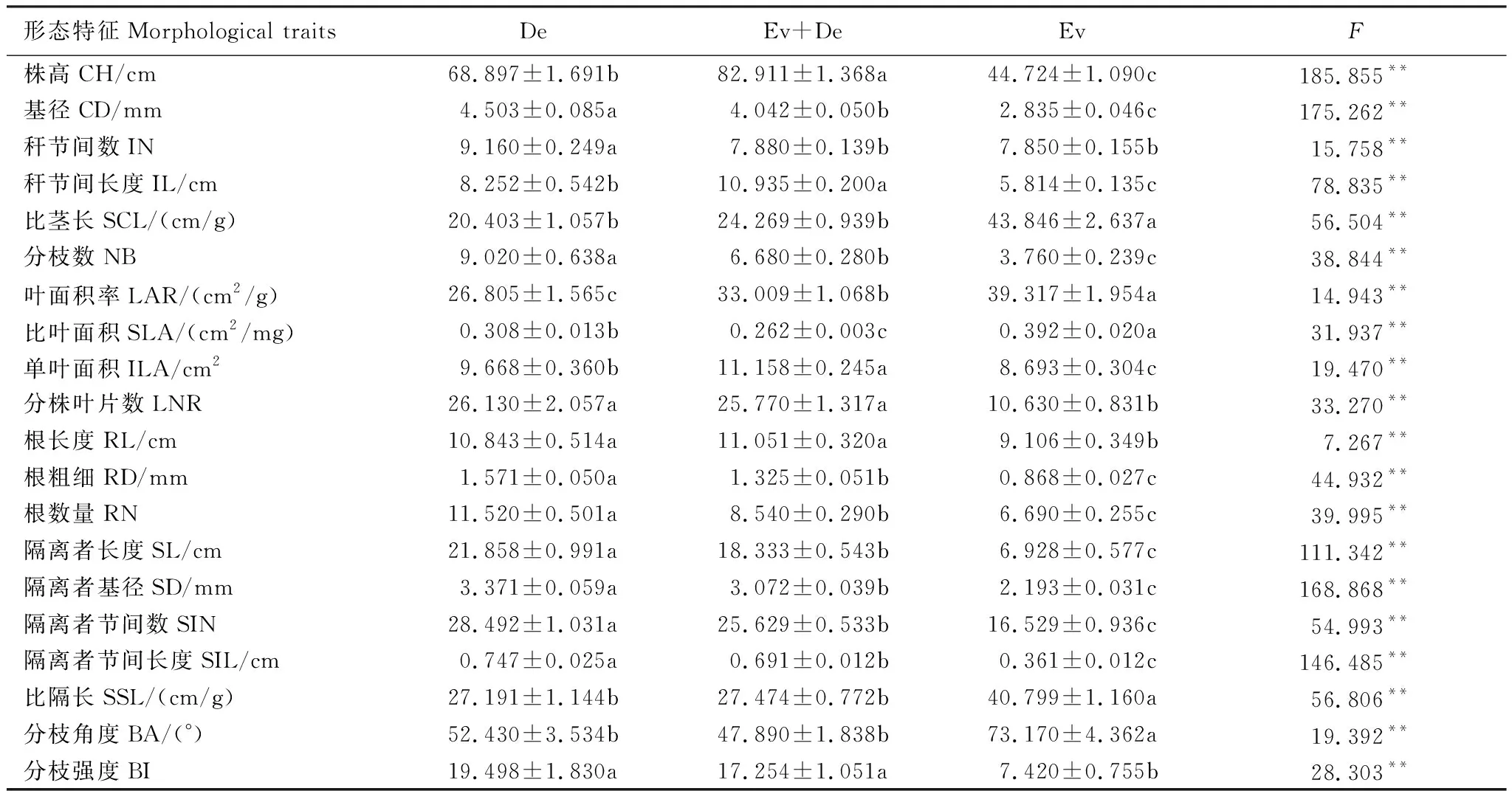
表3 不同林型下紫耳箭竹分株种群的形态特征(平均值±SE)
CH:株高,Culm height;CD:基径,Culm diameter;IN:秆节间数,Internode number;IL:秆节间长度,Internode length;SCL:比茎长,Specific culm length;NB:分枝数,Number of branch;LAR:叶面积率,Leaf area ratio;SLA:比叶面积,Specific leaf area;ILA:单叶面积,Single leaf area;LNR:分株叶片数,Leaf number per ramet;RL:根长度,Root length;RD:根粗细,Root diameter;RN:根数量,Root number;SL:隔离者长度,Spacer length;SD:隔离者基径,Spacer diamater;SIN:隔离者节间数,Spacer internode number;SIL:隔离者节间长度,Spacer internode length;SSL:比隔长,Specific spacer length;BA:分枝角度,Branch angle of spacer;BI:分枝强度,Branch intensity of spacer;同行不同字母表示差异显著;*P<0.05;**P<0.01
3.3 紫耳箭竹分株种群的生物量积累和生物量分配特征
不同冠层结构及其光环境对紫耳箭竹分株种群的生物量积累和生物量分配特征均影响显著(P<0.05)。常绿阔叶林下,紫耳箭竹分株种群各构件生物量(秆,枝,叶,隔离者,根)、地上生物量和总生物量均显著低于其他两个林型(P<0.05);而在落叶阔叶林和混交林中,紫耳箭竹分株种群的秆生物量、隔离者生物量、根生物量、地上生物量和总生物量无显著差异(P>0.05),但枝和叶生物量表现为:混交林>落叶阔叶林(P<0.05)(图2)。
生物量分配特征中,不同林型下的紫耳箭竹秆生物量占的比例均为最大,超过50%,根生物量占的比例均为最小。在落叶阔叶林中,紫耳箭竹种群分配较多的生物量用于隔离者的生长,而减少对枝、叶生物量的分配(P<0.05);混交林中,紫耳箭竹分配较多的生物量用于枝的侧向生长(P<0.05);常绿阔叶林中,紫耳箭竹则分配较多的生物量用于秆和根系的生长,而降低对枝、隔离者生物量的分配(P<0.05)。从总体分配情况来看,混交林下生长的紫耳箭竹种群将较多的生物量用于地上部分的生长;相反,落叶阔叶林下的则将较多的生物量用于地下部分的生长(P<0.05),但其与常绿阔叶林中的无显著差异(P>0.05)(图2)。

图2 不同林型下紫耳箭分株生物量积累(A)和生物量分配(B)特征(平均值±SE)Fig.2 The biomass and biomass allocation of F. decurvata ramets under different forest types (Means ± SE)CM:秆生物量,Culm biomass;BM:枝生物量,Branch biomass;LM:叶生物量,Leaf biomass;SM:隔离者生物量,Spacer biomass;RM:根生物量,Root biomass;AM:地上生物量,Above-ground biomass;TM:总生物量,Total ramet biomass;CP:秆生物量百分率,Allocation percentage to culm;BP:枝生物量百分率,Allocation percentage to branch;LP:叶片生物量百分率,Allocation percentage to leaf;SP:隔离者生物量百分率,Allocation percentage to spacer;RP:根生物量百分率,Allocation percentage to root;AP:地上生物量百分率,Allocation percentage to above-ground;图中同一性状不同字母表示差异显著
3.4 紫耳箭竹分株种群性状特征的可塑性指数
为了比较紫耳箭竹分株种群的各性状特征对冠层结构改变敏感度的差异,对其各性状特征的可塑性指数进行了排序(图3)。结果表明,紫耳箭竹可塑性指数最大的性状为枝干重BM(0.816),可塑性指数最小的性状为地上生物量百分率AP(0.034)。紫耳箭竹的生物量累积特征可塑性指数平均值、形态特征可塑性指数平均值、生物量分配特征可塑性指数平均值分别为0.816,0.683,0.419,也就是说紫耳箭竹以其生物量累积特征对冠层结构改变的敏感度最大,形态特征次之,生物量分配特征的敏感度最差。

图3 紫耳箭竹分株种群性状特征的可塑性指数Fig.3 Plasticity index of F. decurvata ramet traits
3.5 紫耳箭竹性状特征与冠层结构和光环境特征的关系
冠层结构和光环境特征对紫耳箭竹性状特征有显著的影响(P<0.05)(表4)。紫耳箭竹种群的分株密度、株高和盖度随着冠层开度、叶面积指数和林下总光照的增加而呈现出先增加,后降低的趋势;但随着平均叶倾角的增加,呈现出先降低,后增加的趋势,并且均在混交林中达到最大值。

表4 紫耳箭竹性状特征与冠层结构和光环境特征的关系
CO:冠层开度,Canopy openness;LAI:叶面积指数,Leaf area index;MLA:平均叶倾角,Mean leaf angle;Tot:林下总光照,Total solar radiation under canopy;RD:分株密度,Ramet density;CH:株高,Culm height;CR:盖度,Coverage;*P<0.05;**P<0.01
4 结论与讨论
通过比较分析金佛山3种典型群落类型的冠层结构特征可知,随着落叶阔叶林常绿落叶阔叶混交林常绿阔叶林演替的进行,群落的冠层结构发生了改变,即群落的冠层开度降低,叶片覆盖率增加,叶倾角变小,趋于水平化,使得冠层对光的截获能力提高,阳光透过冠层到达下方的辐射量越少,透光率减小(图1),而林下光资源是克隆植物发生形态可塑性变化的重要诱因[33]。在异质性强的生境内,克隆植物能够通过调节形态特征、生物量积累特征和生物量分配特征来适应不同的光照水平,进而达到在不同生境中扩展生存的目的。结果表明,紫耳箭竹种群形态特征的可塑性指数较大,对光环境改变敏感(图3)。表现为在光照水平低的常绿阔叶林中,紫耳箭竹的株高、基径、节间数、节间长度、分枝数、单叶面积、隔离者特征(隔离者长度、直径、节间长度)和根特征(根粗细和数量)均达到最小值(表3)。当土壤水分充足时,林内光照强度的降低会导致分株光合产物(总生物量)积累下降,从而限制植株的高生长[16, 34- 35],而地下茎能够对资源性质和资源量做出反应,将较多的分株选择性地集中在地下资源相对丰富的小生境内[36],所以地下茎短小;根系则对丰富的地下资源水平也没表现出较强的觅食行为[37- 38]。但是在该低光环境下,紫耳箭竹的比叶面积、叶面积率显著增大,从而提高分株的光能利用效率;比茎长也显著增大,说明单位生物量的茎杆在低光环境中有长得更长的趋势;而分枝角度和比隔长也显著大于其他两个林型,则有利于分株在郁闭林冠下的分散生长,是克隆植物为适应不同光环境而发生的可塑性变化[39- 41],这与亚高山暗针叶林林窗环境中华西箭竹(Fargesianitida)的研究结果相似[17- 19, 23]。在光照条件最好的落叶阔叶林中,紫耳箭竹种群产生了长而粗的地下茎,使得分株较为分散,更有效地对土壤水分觅食,以便搜索并且获取必须的资源,这是它克隆生长的一种适应策略[42],并且通过增大根的吸收面积和数量,以能够提高对水的吸收效率,适应生存[37- 38]。而分枝强度随光资源水平的上升而增加,表明克隆植物利用资源的能力以及产生无性分株的能力增强[43]。但是紫耳箭竹种群的株高、秆节间长度、单叶叶面积并非在该林型内最大,而是在混交林内。相关回归分析的结果也表明,冠层结构和光环境显著影响着紫耳箭竹的性状特征。紫耳箭竹的分株密度、株高和盖度也是在混交林中达到最大值(表4)。已有研究表明光照强度的增加会快土壤水分散失,造成水资源贫乏,因此植物把更多的生物量分配到地下部分,把分株放置在较远的地方,通过隔离者的运输来实现资源共享[16],同时减少地上部分的生长,从而利于对水分资源的获取[44- 45],可认为紫耳箭竹对变化的光环境可能存在行为表现和生存策略的差异[46]。
植物的生物量分配格局是适应环境的一种生存策略[47],又反应了环境中可利用资源水平的变化[48]。在不同的环境条件下,植物总是对生长、繁殖和维持等功能间进行资源分配的权衡(trade-off),使其各功能分配达到最佳状态[49]。因此,生物量分配格局可塑性具有重要的生态适应意义[50]。本研究中,紫耳箭竹生物量积累特征的可塑性指数最大,敏感度最强。在光照水平最差的林型中,紫耳箭竹分株积累的生物量最少(图2A),主要是由于林分冠层郁闭大,透过冠层进入林内的光照相应减少,叶片进行光合作用的能力减弱,但其仍然要消耗部分养分,故造成单个克隆的生物量减小[51]。生物量分配特征的可塑性指数虽然最小,但仍对光环境的改变有重要的指示作用。因此,在该光照水平差的林冠环境下,紫耳箭竹通过加大对光合构件——叶,以及支持构件——秆的生物量的投资,以期提高对光资源的获取能力,并减少对地下茎生物量的分配(图2B),从而导致低光环境下产生的无性系分株减少,繁殖能力降低[44]。随着常绿阔叶林常绿落叶阔叶混交林落叶阔叶林下光照强度的增加,紫耳箭竹对隔离者生物量的投资呈现增大的趋势。表明当光照充足而地下资源水平低时,能量主要投资于资源吸收结构,以便提高资源的吸收[44- 45],这与前面的研究一致。Liu等对斑苦竹(Pleioblastusmaculata)研究也表明,随林冠光照强度的增大,无性系投入较多的资源于地下部分无性系的生长和根部的构建[42, 52]。但本研究中紫耳箭竹对根生物量的投资表现出相反的趋势,在常绿阔叶林下对根系的投资比例反而最大。这种差异表明不同物种的反应不完全一致,克隆植物的觅食行为和生态适应是多样的[43]。
综上所述,紫耳箭竹对变化的冠层结构和光环境的表现出较为敏感的形态可塑性。在光照条件好的环境中,紫耳箭竹生长良好,降低对枝、叶投资,将较多的生物量用于产生长而粗的地下茎,利于对空间资源的获取以及无性系分株的产生;光照条件差时,分株变得矮小,叶片变窄,分枝变少,根系少而短,隔离者短而小,分枝强度降低,繁殖能力降低,但通过增加叶面积率、比叶面积、比茎长、比隔长和分枝角度在郁闭的环境下适应生存。由此可知,群落的内部可以通过调控冠层结构及其光环境的改变,影响小径竹的形态可塑性,从而可以达到协调和控制小径竹种群的发展。
猜你喜欢
作文成功之路·教育教学研究(2024年1期)2024-04-09 17:58:36
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
绿色中国(2019年18期)2020-01-04 01:57:08
南方农业学报(2018年4期)2018-09-10 07:22:44
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
西南医科大学学报(2015年1期)2015-08-22 13:01:48
西部林业科学(2015年3期)2015-01-17 05:42:50
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:55
