
盐旱交叉胁迫对灰胡杨(Populus pruinosa)幼苗生长和生理生化特性的影响
2018-11-14 12:46:36王利界周智彬范敬龙范文鹏
生态学报 2018年19期
王利界,周智彬,常 青,范敬龙,范文鹏
1 中国科学院新疆生态与地理研究所,乌鲁木齐 830011 2 四川师范大学地理与资源科学学院,成都 610101 3 中国科学院大学,北京 100049
干旱和盐胁迫不仅影响植物的生长发育,而且也影响植物的各种生理代谢活动[1- 2]。我国大约有3300 hm2的盐渍地面积,干旱、半干旱地区分布在华北、西北内蒙和新疆等地区,其面积约占全国土地面积的1/2,干旱、沙漠化日益加剧,已经成为我国面临的重大环境问题[3]。植物在受到干旱及盐胁迫时会产生复杂的逆境响应过程[4- 5],研究干旱及盐胁迫对植物生长和生理特性的影响对理解植物抗旱抗盐机理具有重要意义。
灰胡杨(Populuspruinosa)属于杨柳科杨属的高大落叶乔木、稀灌木[6],主要分布在塔里木河流域,具有防风固沙、稳定河道、保护绿洲和维持荒漠区脆弱环境生态平衡的功能[7]。目前对灰胡杨的研究多集中在盐分或水分等单一胁迫。如:王海珍在不同干旱胁迫下对灰胡杨幼苗进行试验,认为灰胡杨幼苗通过积累大量游离的脯氨酸(Pro)来提高抗旱性[8];在土壤水分条件较好时,灰胡杨蒸腾耗水量高,水分利用比较充分[9];胡利华通过对盐胁迫下灰叶胡杨幼苗的初步研究,得出在200 mmol/L的盐分处理下,灰叶胡杨的愈伤组织表现出快速的生长[10];李志军采用盆栽试验研究在不同干旱胁迫下灰胡杨的光合作用发现,在干旱胁迫进程中轻度和重度干旱胁迫下灰胡杨净光合作用速率经历了急剧下降—缓慢回升—下降3个阶段,在干旱胁迫下PSII活性下降明显,随着胁迫时间的延长,FV/Fm和FV/Fo的下降幅度增加[11- 12]。逆境下植物能够感应外界胁迫,并能通过自身的调节系统,使之在生理和分布上发生适应性反应,以增强在胁迫条件下的生存机会。本文以1年生灰胡杨幼苗为研究对象,通过对灰胡杨在盐旱交叉胁迫条件下保护酶系统的研究,旨在揭示其耐盐耐旱机理,为沙漠腹地引种优良植物品种提供理论基础。
1 研究区概况
研究区位于塔克拉玛干沙漠腹地的塔克拉玛干沙漠研究站(塔中沙漠植物园),地理坐标为39°01′N,83°36′E,海拔在1100 m左右,据塔中气象站资料,该地区年均气温12.4℃,最热月7月平均气温28.2℃,沙面最高温度可达75.3℃,最冷月12月平均气温-8.1℃,极端最高气温45.6℃,极端最低气温-22.2℃,差为67.8℃;年平均降水量24.6 mm,平均相对湿度29.4%,年潜在蒸发量3639 mm;年平均风速2.5 m/s,最大瞬时风速20 m/s。植物园内从各地引种种植了多种能够适应干旱环境的植物,其中以多种属的怪柳、梭梭和沙拐枣为主,塔中植物园利用矿化度为4.04 g/L的地下咸水灌溉,灌水量为30 L,灌溉周期平均为10 d。
2 材料与方法
2.1 试验材料
本实验供试材料为1年生灰胡杨幼苗,2015年9月在新疆阿克苏选取生长状态一致的自然实生幼苗,移栽至试验田中,缓苗至次年6月。
2.2 试验方法
试验采用随机区组实验设计(表1),包括1个对照,3个单一盐分胁迫,2个干旱胁迫以及6个盐旱交互胁迫,其中3个盐分胁迫处理为B(8 g/L),C(11 g/L),D(15 g/L)。2个干旱胁迫处理的梯度为灌水量T1(25 L)和T2(15 L),使土壤相对含水量符合Hsiao[13]提出的植物水梯度划分设计。6个盐旱交互为B,C,D和T1,T2随机组合,每个处理设5个重复。试验区四周埋设3层防渗土工膜隔离,防渗膜埋深1.2 m,用以防止小区之间灌溉水分和盐分的相互渗透对实验的影响。2015年11月至2016年5月缓苗期间,采用矿化度水平约为4 g/L的地下水充分灌水。待苗木生长完全恢复正常后,于2016年5月30日开始咸水滴灌控制实验,灌水周期为12 d,灌水量维持在设定梯度。咸水灌溉处理持续至2016年9月15日结束。

表1 盐旱胁迫试验设计
3个盐分胁迫处理为B(8 g/L),C(11 g/L),D(15 g/L)。2个干旱胁迫处理的梯度为灌水量T1(25 L)和T2(15 L)
2.3 测定方法
2.3.1 生长指标的测定
每个灌水周期结束后的一天,用卷尺测量植株高度、东西最长冠径和南北最长冠径以及标定枝长。
2.3.2 生理生化指标的测定
试验结束取样时,每个处理取样3份,分别测定其相关指标。超氧化物歧化酶(SOD)活性采用氯化硝基四氮唑蓝(NBT)还原法[14];过氧化物酶(POD)活性采用愈创木酚法[15];过氧化氢酶(CAT)活性采用可见光法[16];丙二醛(MDA)含量的测定采用硫代巴比妥酸法[17];游离的脯氨酸(Pro)含量的测定采用酸性茚三酮显色法[18]测定。
2.4 数据处理
用DPS 7.05软件进行方差分析,比较处理间的差异显著性,采用Origin软件制图。
3 结果分析
3.1 不同灌水量、盐分和交互作用下对灰胡杨幼苗生长的影响
植物的表观生长能够比较直观的反应植物所在环境的优劣,生长抑制是植物对逆境响应最敏感的过程[19]。如表2所示,在单一的干旱或盐胁迫下,随着灌水量的减少或盐浓度的增加,冠幅和枝长的增量受到了明显的抑制,降低的幅度分别为25.0%、37.0%(干旱胁迫)、71.0%、88.09%(盐胁迫)差异显著(P<0.05)。干旱胁迫对株高的影响并不大,而株高增长量随着盐胁迫的加重,在B处理下先升高,随后在C、D下逐渐降低,差异显著(P<0.05),表明灰胡杨具有一定的耐旱性。说明在单一的干旱和盐胁迫下,灰胡杨受到盐胁迫的影响显著大于干旱。
表2不同灌水量、盐浓度及交叉胁迫下对灰胡杨幼苗株高、冠幅和枝长增长量的影响
Table2EffectsofdifferentirrigationamountandsaltconcentrationandintercrossstressonincreaseofplantheightandcrowndiameterandbranchlengthinPopuluspruinosa
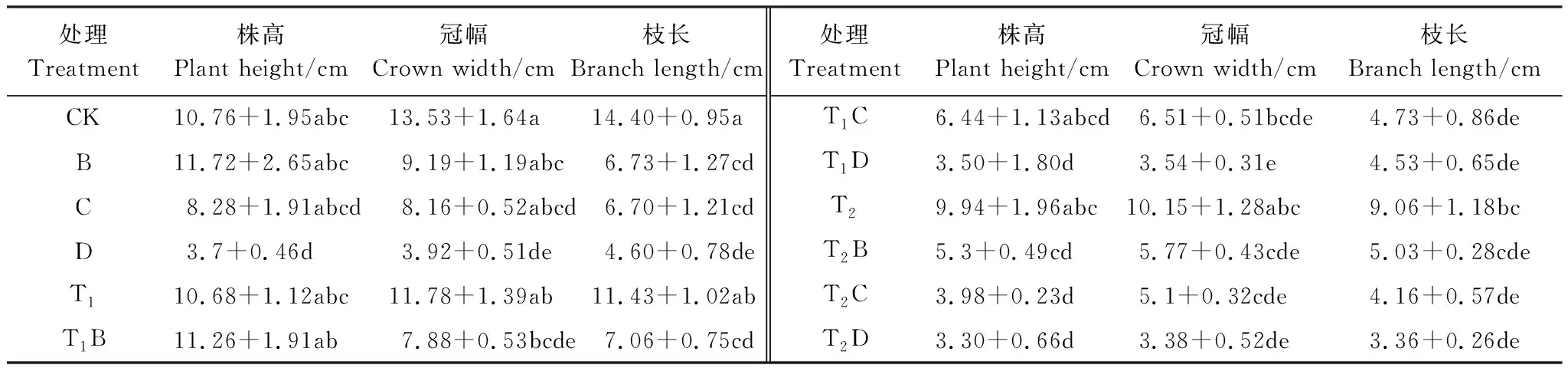
处理Treatment株高Plant height/cm冠幅Crown width/cm枝长Branch length/cm处理Treatment株高Plant height/cm冠幅Crown width/cm枝长Branch length/cmCK10.76+1.95abc13.53+1.64a14.40+0.95aT1C6.44+1.13abcd6.51+0.51bcde4.73+0.86deB11.72+2.65abc9.19+1.19abc6.73+1.27cdT1D3.50+1.80d3.54+0.31e4.53+0.65deC8.28+1.91abcd8.16+0.52abcd6.70+1.21cdT29.94+1.96abc10.15+1.28abc9.06+1.18bcD3.7+0.46d3.92+0.51de4.60+0.78deT2B5.3+0.49cd5.77+0.43cde5.03+0.28cdeT110.68+1.12abc11.78+1.39ab11.43+1.02abT2C3.98+0.23d5.1+0.32cde4.16+0.57deT1B11.26+1.91ab7.88+0.53bcde7.06+0.75cdT2D3.30+0.66d3.38+0.52de3.36+0.26de
相同的字母表示各处理之间差异不显著;不同的字母表示差异显著(P<0.05)
在水盐交互胁迫下,灰胡杨生长较单因子胁迫下要复杂。不同的交叉胁迫处理,随着胁迫程度的增加,灰胡杨冠幅和枝长增长量显著降低。T1和不同盐浓度交互胁迫下与对照相比,株高、冠幅和枝长的最大降幅分别为67.5%;73.8%和68.5%。T2和不同盐浓度交互胁迫下与对照相比,株高、冠幅和枝长的最大降幅分别为69.3%;75%和76.6%。降幅相互之间并没有差异,说明在盐旱交互胁迫下,盐浓度是影响灰胡杨幼苗生长的主要因子。同样,双因素方差分析表明,在灌水量、盐分以及盐旱交互处理下,只有盐分胁迫对灰胡杨相对高生长(F水=2.679,F盐=6.238,F水+盐=0.722)、相对枝长(F水=2.18,F盐=20.252,F水+盐=1.389)以及冠幅增量(F水=1.912,F盐=9.779,F水+盐=0.141)产生了显著影响(P<0.05),而不同的灌水量、水盐交互处理均没有显著影响。
3.2 不同灌水量、盐分和交互作用下对灰胡杨幼苗生理生化影响
3.2.1 不同灌水量、盐分和交互作用下对灰胡杨幼苗叶片超氧化物歧化酶活性的影响
在逆境胁迫下,木本植物可动员酶性和非酶性两类防御系统保护细胞免遭氧化伤害,前者包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT);后者包括还原型谷胱甘肽,类胡萝卜素(CAR)等。SOD、POD、CAT是植物体内重要的活性氧清除酶,其中SOD它是氧自由基代谢的第一个酶类,被认为是关键酶。
如图1和表3可知,在干旱胁迫下,灰胡杨幼苗SOD 活性随着干旱胁迫程度的增加逐渐增加,T1与CK、T2与T1相比,增幅分别为7%、2%,差异不显著,但T2与CK相比增幅为9%,差异显著(P<0.05)。说明灰胡杨对于干旱胁迫具有良好的适应能力。在盐胁迫下灰胡杨幼苗SOD活性随着胁迫程度的加重,呈现先上升后下降的趋势,在盐浓度B处理下增幅达到最大,为29.1%,与对照存在显著差异(P<0.05)。在D处理下SOD活性相比对照有所下降,说明在较高浓度的盐胁迫下,SOD活性已经受到抑制,无法维持正常水平。表明在单一的盐、旱胁迫下,灰胡杨抗旱能力要大于抗盐能力。在盐旱交互胁迫下,除了T2D处理,灰胡杨幼苗SOD活性均上升,且与对照存在显著差异,各处理间也存在显著差异(P<0.05)。在T1和不同盐分交互胁迫下,随着胁迫的加重,SOD活性先增高达到峰值随后降低,与对照相比增幅分别为43.3%、31.1%、1.5%。在T2处理和不同盐分交互胁迫下,随着胁迫的加重,SOD活性先增高后降低,在T2D处理下,相比对照SOD活性有所下降,降幅为24.7%。说明灰胡杨在水盐交互胁迫下,干旱胁迫占据主导地位。在适度的盐旱交互胁迫下,可以诱导或激活SOD,增加灰胡杨幼苗的抗逆性,但是重度的盐旱交互胁迫会严重打破灰胡杨自由基产生和清除之间的平衡,严重破坏了膜透性,从而抑制灰胡杨的生长。
表3不同灌水量不同盐含量下灰胡杨生化特征重复测量结果的方差分析
Table3RepeatedmeasuresANOVAontheeffectsofirrigationamountandsaltconcentrationonphysiologicalandbiochemicalcharacteristicsinPopuluspruinosa
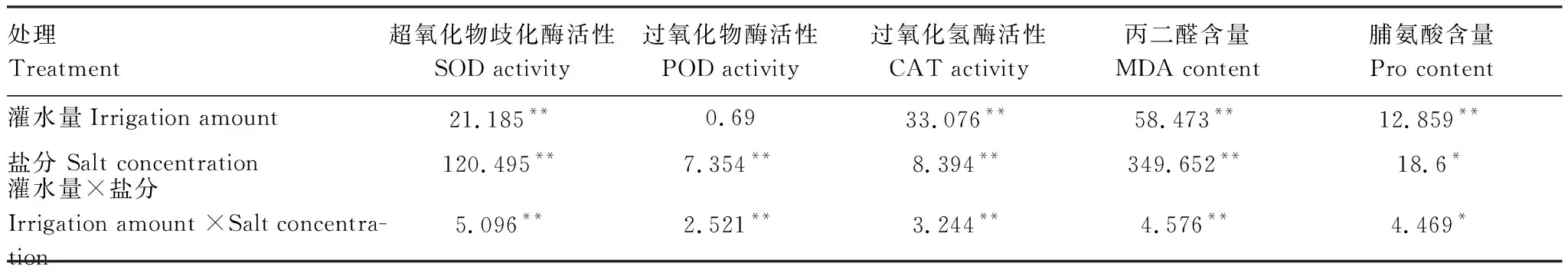
处理Treatment超氧化物歧化酶活性SOD activity过氧化物酶活性POD activity过氧化氢酶活性CAT activity丙二醛含量MDA content脯氨酸含量Pro content灌水量Irrigation amount21.185**0.6933.076**58.473**12.859**盐分 Salt concentration120.495**7.354**8.394**349.652**18.6*灌水量×盐分 Irrigation amount ×Salt concentra-tion5.096**2.521**3.244**4.576**4.469*
SOD, superoxide; POD, peroxidase; CAT, catalase; MDA, malonyldialdehyde; Pro, proline. *,P<0.05; **,P<0.01
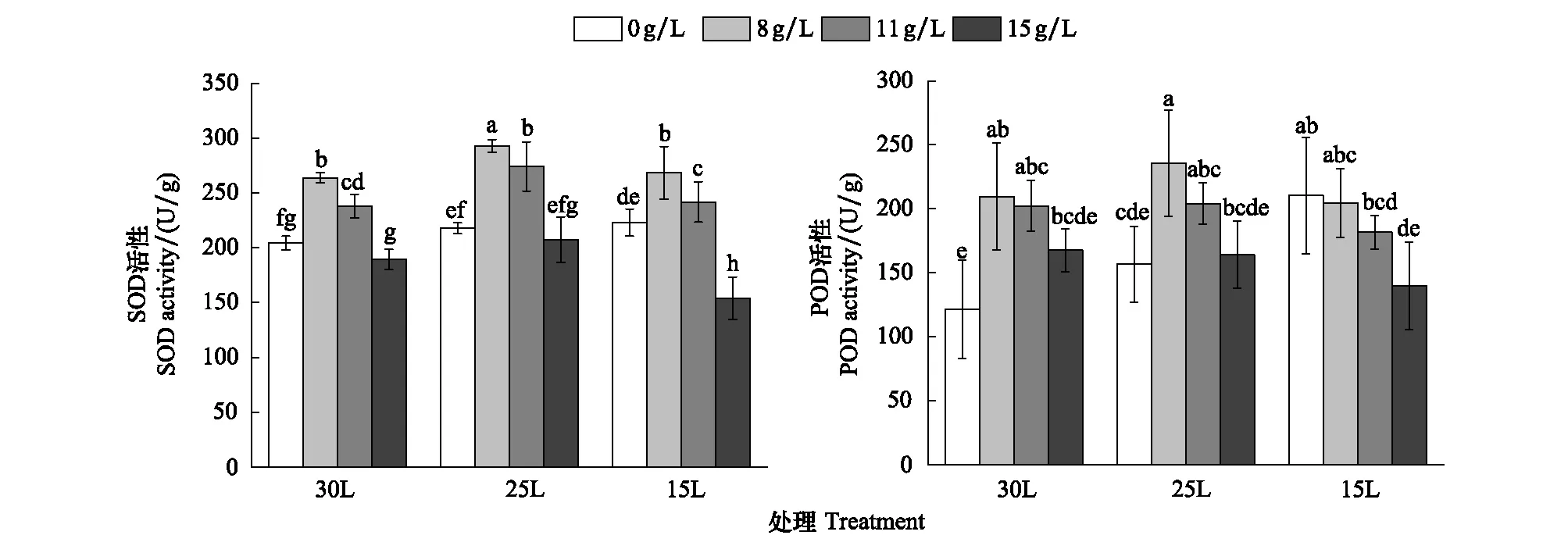
图1 灰胡杨不同盐旱胁迫下SOD和POD活性的变化Fig.1 Changes of SOD and POD activities under different irrigation amount and salt concentration in Populus pruinosaSOD: 超氧化物歧化酶, superoxide; POD: 过氧化物酶, peroxidase
3.2.2 不同灌水量、盐分和交互作用下对灰胡杨幼苗叶片过氧化物酶活性的影响
如图1和表3可知,干旱胁迫下灰胡杨幼苗POD活性随着胁迫程度的加重,呈现先上升的趋势,且与对照有显著差异(P<0.05),在T1、T2处理下,增幅分别为29.1%、73.4%。说明灰胡杨幼苗的POD对其在干旱胁迫下自由基的清除具有重要作用。在盐胁迫下,叶胡杨幼苗POD活性随着胁迫程度的加重,呈现先增加后降低的趋势。在B处理下POD活性达到最高,与对照的差异达到显著水平(P<0.05),增幅为72.8%,说明低浓度的盐胁迫,可以激活灰胡杨幼苗POD活性,增强其抗旱能力。随着盐浓度的继续增加,POD活性显著降低,在D处理下,达到最低,但仍高于对照,增幅为38.2%,表明高浓度的盐会抑制POD活性,使其不能维持正常水平,生长受到抑制,这也说明灰胡杨幼苗抗旱性要强于抗盐性。对于盐旱交互胁迫来说,灰胡杨幼苗POD活性呈现先增加后下降的趋势,且与对照存在显著差异。在T1与不同浓度盐分交互胁迫下,POD活性增幅分别为94.2%、68.1%、35.2%,在T2D处理下,POD活性增幅达到最小,为15.4%,与对照相比有显著差异(P<0.05)。已上分析表明在,盐旱交互胁迫下,灰胡杨幼苗受到刺激,POD活性升高,抗逆性增强,但随着胁迫程度的加重,POD活性显著降低,却仍然高于对照,但差异并不显著(P>0.05),此时,灰胡杨幼苗POD仍然发挥着作用,由此我们推断,若盐旱交互胁迫进一步加重,超过试验所设处理程度,则灰胡杨幼苗POD活性会受到抑制。

图2 灰胡杨不同盐旱胁迫下CAT活性和MDA含量的变化Fig.2 Changes of CAT activities and MDA under different irrigation amount and salt concentration in Populus pruinosaCAT: 过氧化氢酶; catalase; MDA: 丙二醛; malonyldialdehyde
CAT主要分布于过氧化氢酶体中,将高浓度的 H2O2清除,与SOD协同反应,使活性氧维持在较低的水平,减轻有毒物质对生物细胞的毒害,推迟或阻碍细胞结构的破坏,使组织保持活力。
图2表明,在干旱胁迫下,灰胡杨幼苗CAT活性随着灌水量的减少逐渐升高,在T1、T2处理下与对照相比,增幅分别为31.3%、72.5%,且差异显著(P<0.05)。在盐胁迫下,灰胡杨幼苗CAT活性随着盐浓度的增加,呈现出先增加后降低的趋势,但仍高于对照并与对照有显著差异(P<0.05),在C处理下达到峰值,增幅为47.9%。这说明随着胁迫程度的增加,CAT酶活性增大,从而清除因盐胁迫而产生的自由基,但是当盐胁迫达到一定程度后,酶活性开始受到抑制,自由基无法快速清除,幼苗受到一定伤害;T2处理下的干旱程度并不能抑制灰胡杨CAT酶活性,而D处理下CAT酶活性虽然显著高于对照,但已经有下降的趋势,说明在单一盐或者干旱胁迫下,灰胡杨幼苗CAT抗旱能力要高于抗盐能力。
盐旱交互胁迫时,在T1与不同盐分浓度交互胁迫下,灰胡杨叶片CAT活性,随着胁迫程度的加重,呈现先增后减的趋势,增幅分别为48.1%,60.6%,29.1%。在T2与不同盐分浓度交互胁迫下,灰胡杨叶片CAT活性同样呈现先增后减的趋势,增幅达到最大,分别为107.1%,77.4%,48.5%。这说明对于灰胡杨幼苗CAT来说,水盐交互胁迫下干旱胁迫占主导地位。
丙二醛(MDA)是脂质过氧化的主要产物之一。它可与细胞膜上的蛋白质、酶等结合、交联使之失活,从而破坏生物膜的结构与功能,对细胞有毒性的物质,对许多生物大分子均有破坏作用,因此MDA可以作为判断膜脂过氧化作用的一种主要指标[20]。MDA也是膜质过氧化作用的最终产物,膜系统受伤害的重要标志之一[21],MDA积累越多表明组织的保护能力越弱[22]。
由图2可知,在单一的盐分或干旱胁迫下,随着胁迫程度的加重,灰胡杨幼苗MDA含量均表现出先降低后增加的趋势,与对照有显著差异(P<0.05),这与抗氧化酶活性先升高后降低的趋势相对应。
盐旱交互胁迫下,在T1与不同盐分浓度交互胁迫下,随着交互胁迫程度的加重,灰胡杨幼苗MDA含量表现出先降低后增加的趋势,但在T2与不同盐分浓度交互胁迫下,灰胡杨幼苗MDA含量却持续增加,差异显著(P<0.05),增幅分别达到23.3%,96.3%,140.6%。
说明在盐旱交互胁迫过程中,随着胁迫程度的加重,灰胡杨幼苗细胞内抗氧化酶活性下降,自由基的平衡已经无法控制,从而引发了脂膜过氧化作用,对细胞产生毒害作用。

图3 灰胡杨不同盐旱胁迫下Pro含量的变化 Fig.3 Changes of Pro activities under different irrigation amount and salt concentration in Populus pruinosaPro: 脯氨酸, proline
脯氨酸是水溶性较大的氨基酸,具有较强的水合力,当植物受到逆境胁迫时,脯氨酸的增加有助于细胞或组织持水[23]。由图3和表3可知,在单一干旱或盐分胁迫下,灰胡杨叶片脯氨酸含量均呈上升趋势,但是盐胁迫下脯氨酸含量增幅显著高于干旱胁迫,且T1与T2相比,脯氨酸含量并无显著差异(P>0.05)。这说明,灰胡杨幼苗在面对单一盐分或干旱胁迫时,脯氨酸的作用更偏向于缓解盐胁迫所带来的损伤。
在盐旱交互胁迫下,随着胁迫的加重,灰胡杨幼苗叶片脯氨酸含量也显著增加,各盐旱交互胁迫处理下的灰胡杨脯氨酸含量均高于各单一盐胁迫或干旱胁迫下的含量且显著差异(P<0.05)。与T1相比,T2和不同盐浓度交互胁迫下,灰胡杨幼苗叶片脯氨酸含量增幅较大,分别为155.3%,188.0%,235.9%。说明随着盐旱交互胁迫程度的加重,脯氨酸含量也持续增加,灰胡杨幼苗抗逆性进一步得到增强。
4 讨论与结论
生长量是对胁迫响应的综合表现,也是反映植物抗旱性和耐盐性的基本标准。在水、盐胁迫下,根系首先感应并发出信号,同时根系形态结构、生理化学生物量等也发生相应变化,进而抑制植物生长,甚至导致植物死亡[24]。有研究表明,落羽杉[25]和金叶白蜡[26]株高,基径和生物增量随着土壤盐分浓度的增加和土壤水分含量的减少而减少。本实验中,通过灌水量和盐浓度的改变,模拟塔克拉玛干沙漠环境,结果表明,随着干旱程度的加重和盐浓度的升高,灰胡杨株高,枝长,冠幅均受到不同程度的抑制,差异显著,这与上述研究结果一致。在盐旱胁迫下,由于外界渗透势过低,灰胡杨无法吸取足够水分,从而影响各种生理反应,导致灰胡杨生长受到抑制,但是两者的胁迫损伤程度并不相同,盐胁迫对植物产生的生长抑制效果明显大于干旱胁迫,干旱胁迫对植物的危害主要是通过渗透胁迫,而盐分对植物的危害主要是通过渗透胁迫和离子毒害[27]。
植物在各种逆境胁迫下,其保护酶活性会发生不一样的变化,在盐旱交互胁迫下,植物叶片内会积累大量的OH-和H2O2,同时体内抗氧化酶SOD、POD、CAT活性增加,及时清除体内有害的活性氧,防止自由基对细胞的伤害,以提高植物的适应能力[28- 29]。庄伟伟[30]通过对银沙槐幼苗在水盐交叉胁迫下的研究表明,在轻度干旱、中度干旱和盐分的单一胁迫或交叉胁迫下银沙槐幼苗保护酶(SOD、POD、CAT)活性均上升,但在重度盐旱交叉胁迫下,保护酶的活性不能维持较高的水平。杨国会[31]通过对甘草的研究表明,随着NaCl浓度的增加,POD、SOD 2种保护酶的活性呈现先上升后下降的趋势。
本研究中灰胡杨幼苗抗氧化酶SOD、POD、CAT活性仅受到干旱胁迫时,随着胁迫的加重,其活性表现出持续增加。而在盐分胁迫和盐旱交叉胁迫下则表现出先升高后下降的趋势,表明灰胡杨幼苗的耐胁迫具有一定的范围,在盐分和灌水量的交互胁迫下,灰胡杨幼苗体内的抗氧化酶系统和具有抗性特征的生理活动被诱导加快,抗氧化酶SOD、POD、CAT活性在此诱导下逐渐增加,以消除盐分和干旱胁迫所导致的叶片中产生过多的活性氧,但是随着胁迫程度的增加,叶片中活性氧数量也在增加,达到某一数量超出正常范围内酶系统所能承受的能力,导致其不足以清除体内的过多自由基,从而造成了细胞膜及酶系统等的破坏,说明保护酶活性是维持在一定胁迫程度之内发生的,超出这一程度后,保护酶SOD、POD、CAT活性会急剧下降。
植物生长环境受到破坏时,膜脂发生过氧化,细胞膜的正常功能受到破坏而使膜与壁分离[32]。MDA是膜脂过氧化过程的终产物,在细胞中积累的多少可以判断过氧化的程度,积累量越多,胞膜系统受伤害的程度越大。赵银河等[33]研究一年生皂角和紫荆实生苗结果表明,随着干旱和盐交互胁迫强度的增加,两树种的抗氧化酶SOD、CAT和POD活性均呈现先下降再上升的趋势,活性都高于正常水平下的酶的活性,而且丙二醛含量也均高于正常水平下。本研究中,随着盐胁迫的加重,灰胡杨叶片MDA含量呈先降低后增加的趋势,与上述结论一致。在T1、T2和较低盐浓度CK、B交互胁迫下,灰胡杨叶片MDA含量也呈先降低后增加的趋势,但是与较高盐浓度C、D交互胁迫下,其含量一直稳定增加,差异显著(P<0.05)。说明在轻度的盐旱胁迫下,保护酶能够发挥作用,从而有效防止膜脂过氧化对灰胡杨的伤害,但在重度胁迫下,保护酶活性受到抑制,自由基的清除无法达到平衡,导致了膜系统受到破坏,从而抑制了植物的生长甚至导致其死亡。
游离脯氨酸(Pro)是植物体内的一种重要的调节物质,能够有效的保持体内的水分平衡,较低的溶质水势。当植物处于逆境下生长时,脯氨酸的积累,以防止蛋白质的脱水变性,持体内生长环境,保证水合能力。王利军等[34]研究发现,在水分和盐分胁迫处理下,沙枣脯氨酸含量随着胁迫时间的增加逐渐增加。刘瑞宁等[35]研究发现,在水盐交互胁迫下,叶底珠幼苗脯氨酸含量呈现逐渐上升趋势。本实验中,无论是单一的盐胁迫、干旱胁迫还是盐旱交互胁迫,灰胡杨脯氨酸含量随着胁迫程度的加重而显著增加,说明水分、盐分及水盐交互胁迫对脯氨酸产生了显著影响。脯氨酸的增加,可以提高细胞渗透调节的能力,降低外界胁迫对灰胡杨叶片细胞造成的伤害,从而增强了灰胡杨的抗逆性。
干旱、盐胁迫及交互胁迫对灰胡杨幼苗的生长和生理特性均有一定影响,灰胡杨幼苗能够在盐分8 g/L、灌水量25 L下良好生长,但在盐浓度为11、15 g/L胁迫和15 L灌水量的胁迫以及交互作用下,其生理活动受到显著抑制,体内抗氧化酶活性降低,但仍高于对照。MDA含量与抗氧化酶活性呈负相关,表现出先降低后升高的趋势。同时,作为渗透调节因子的脯氨酸,在整个胁迫试验过程中表现出持续增加的趋势。灰胡杨幼苗通过调节自身生长、体内生理代谢等活动来适应干旱与盐的胁迫,并对胁迫表现出较强的耐受性,从而减少外界对幼苗的伤害。可见,在灌水量为大于30 L,盐分浓度平均为5 g/L的沙漠腹地,灰胡杨可以健康的成长,因此在沙漠腹地可以引种灰胡杨。
猜你喜欢
中老年保健(2022年3期)2022-11-21 09:40:36
草业学报(2022年5期)2022-07-08 06:24:40
金桥(2021年11期)2021-11-20 06:37:30
草业学报(2021年3期)2021-03-22 02:22:00
北广人物(2020年47期)2020-12-09 06:24:12
科学与财富(2019年15期)2019-10-21 13:35:48
草业学报(2018年10期)2018-10-19 05:25:44
发酵科技通讯(2018年2期)2018-07-06 11:38:46
创新作文(1-2年级)(2018年6期)2018-01-22 12:25:50
电镀与环保(2017年5期)2017-12-19 12:06:03
