
云南省贡山地区小型兽类几种细菌性病原体携带现况调查*
2018-11-10 01:53:40李伶俐左曙青杜春红彭红红赵秋敏张久松
寄生虫与医学昆虫学报 2018年1期
李伶俐 左曙青 杜春红 彭红红赵秋敏 代 科 张久松**
(1.安徽医科大学,安徽合肥 230022; 2.军事科学院军事医学研究院微生物流行病研究所,病原微生物生物安全国家重点实验室,媒介生物危害和自然疫源性疾病北京市重点实验室,北京 100071; 3.云南省地方病防治所,云南大理 671000)
云南贡山独龙族怒族自治县位于东经98°08′~98°56′,北纬27°29′~28°23′之间的滇西北怒江大峡谷北段,东与云南省德钦、维西两县相连,西与缅甸毗邻,南与福贡县相邻,北与西藏自治区察隅县接壤。高黎贡山地区物种丰富,素有“世界意义的陆地生物多样性关键地区”之称。小型兽类,特别是啮齿动物作为自然疫源性疾病最重要的宿主可携带和传播多种与疾病相关的病原体(Meerburgetal., 2009;Luisetal., 2013;Palmeirimetal., 2014),如蜱传脑炎病毒、鼠疫耶尔森氏菌、汉坦病毒、钩端螺旋体、莱姆病螺旋体、斑点热立克次体、恙虫病东方体、埃立克体等,并可使人或动物患病,影响生活质量,严重者可致死亡。据报道目前全世界鼠类有35个科389个属,约2 700多种,占世界已知哺乳动物种类数的42%,大约90%的鼠种能携带病原体,有57种与人类疾病相关(郑剑宁等,2007)。我国鼠类(啮齿类)有近200种,已经查明可传播疾病的近80种(琚俊科等,2010)。贡山地区物种丰富,此前在该地区对小型兽类的分布进行调查,调查结果涉及5目12科42属76种鼠形小型兽类(高耀亭等,1962;龚正达等,1989;李璋鸿等,2005)。目前对于云南省自然疫源性疾病的调查主要集中在西南边界地区(吴爱国等,2003;Zhangetal.,2007;张胜勇等,2008;Guoetal.,2013;梁中平等,2014;万道正等,2017),在西北地区的相关报告较少(刘洪光等,2010;Schmidtetal., 2014)。本研究对贡山地区小型兽类病原携带情况进行调查,为自然疫源性疾病的防制提供依据。
1 材料与方法
1.1 标本采集与鉴定
2014—2015年10和11月在云南贡山地区设置采样点,采用夹夜法和笼捕法诱捕野生小型兽类,即调查当天傍晚时分用鼠夹或鼠笼加食饵随机诱捕小兽,次日清晨用特制的布袋收集。将诱捕的小兽在实验室内用乙醚麻醉或机械处死,确定其种属后解剖,取肺、脾标本于液氮中进行保存。
1.2 主要仪器和试剂
研磨组织所用的仪器为美国Next Advance公司生产的Bullet Blender组织细胞破碎仪,PCR扩增所用仪器为美国Applied Biosystems公司生产的GeneAmp 9700 PCR扩增仪,凝胶成像系统来源于法国Viber Loumat公司。DNA聚合酶采用宝生物工程(大连)有限公司生产的Ex-Taq或r-Taq。DNA提取试剂盒采用罗氏High Pure PCR Template Preparation Kit。
1.3 核酸提取与PCR扩增
取20 mg左右的小兽组织研磨匀浆后,12 000 r/min离心3 min,离心后得到的沉淀根据试剂盒说明书进行DNA提取,并将得到的DNA置于-40 ℃保存备用。采用巢式PCR检测无形体Anaplasmataceae(A.p.)、恙虫病东方体Orientiatsutsugamushi(O.t.)、斑点热群立克次体Spotted fever group rickettsiae(SFGR)以及莱姆病螺旋体Lyme disease spirochete等4类病原体。引物根据基因库中不同种属病原体多基因核苷酸序列信息设计或者采用已发表的引物序列,委托北京天一辉远生物科技有限公司合成,引物序列见表1。反应体系为25 μL,其中包括2×Taq Buffer 12.5 μL、10 μmol/L的上下游引物各0.5 μL、模板DNA 4 μL(巢式PCR第2轮模板为0.25 μL第1轮反应产物),加无核酸酶的水补足25 μL。
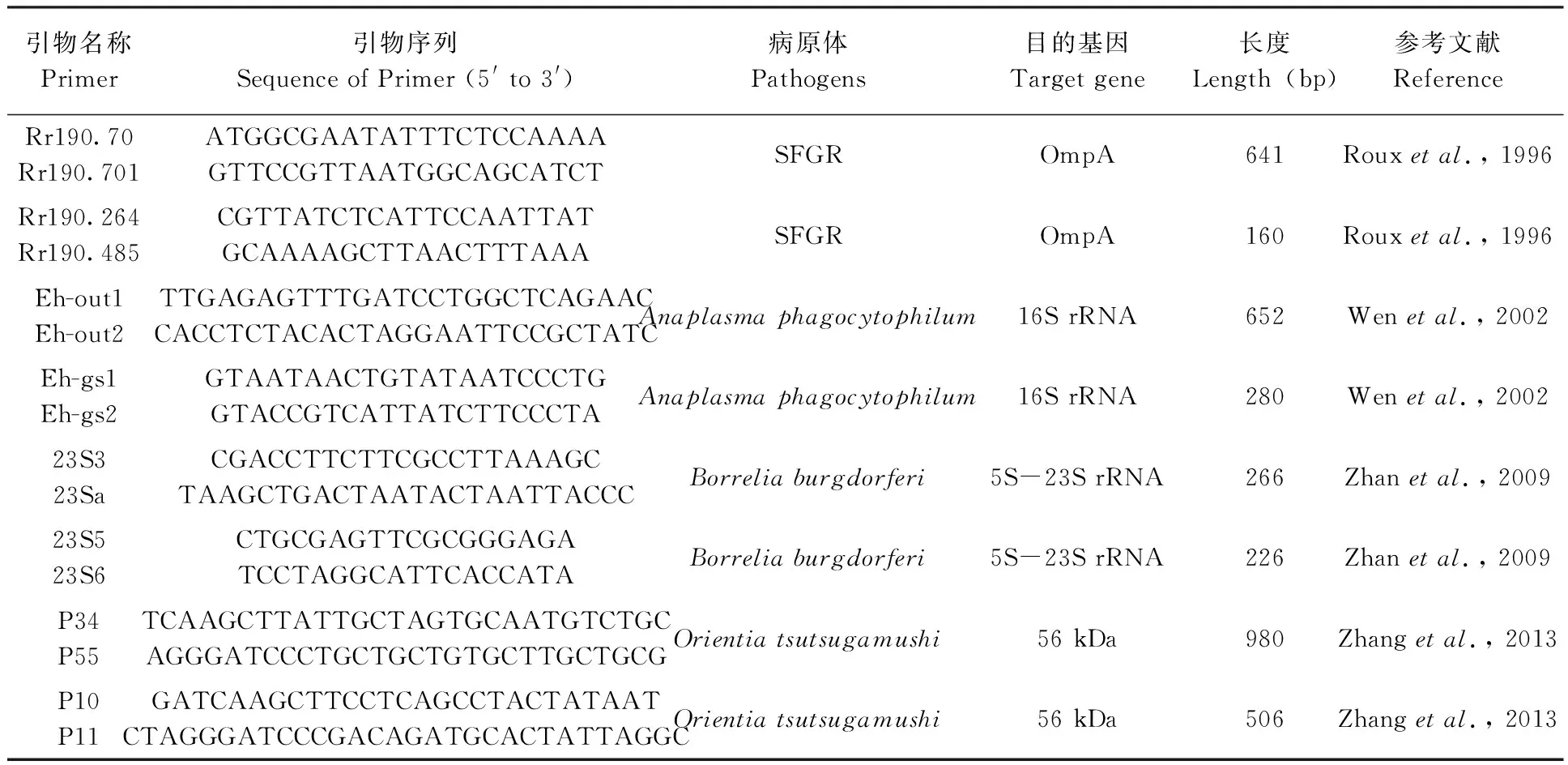
表1 本研究用于病原体检测的引物信息Tab.1 Primers used for detection of bacterial pathogens in this study
1.4 核苷酸序列分析
阳性PCR扩增产物送北京天一辉远生物科技有限公司进行序列测定,将得到的序列信息在GenBank库中进行Blast同源性比对,选择与该序列同源性相对较高的种属代表株或血清型代表株以及标准株进行系统发生和进化分析,利用ClustalX2和Mega 6.0软件进行碱基配对后,用Neighbor-joining方法完成进化树的构建并计算遗传距离,模型采用Kimura-2-parameter model,Bootstrap值设定为1 000。
2 结果
2.1 捕获小型兽类情况
共捕获小型兽类759只,涉及14个属35种。其中构成比大于5%的包括姬鼠属Apodemus221只(29.1 %)、绒鼠属Eothenomys132只(17.4%)、长尾鼩属Soriculus105只(13.8%)、白腹鼠属Niviventer69只(9.1%)、鼩鼱属Sorex58只(7.6%)、短尾鼩属Anourosorex58只(7.6%);构成比低于5%的包括鼠兔属Ochotona27只(3.6%)、家鼠属Rattus24只(3.2%)以及麝鼩属Crocidura、田鼠属Microtus、鼩鼹属Uropsilus、攀鼠属Vernaya、针鼹属Tachyglossus和长吻松鼠属Dremomys。经进一步物种鉴定,该地区野生小型兽类物种构成如图1所示,其中中华姬鼠Apodemusdraco181只,未明确分类的绒鼠Eothenomys80只,分别占23.8%和10.5%。
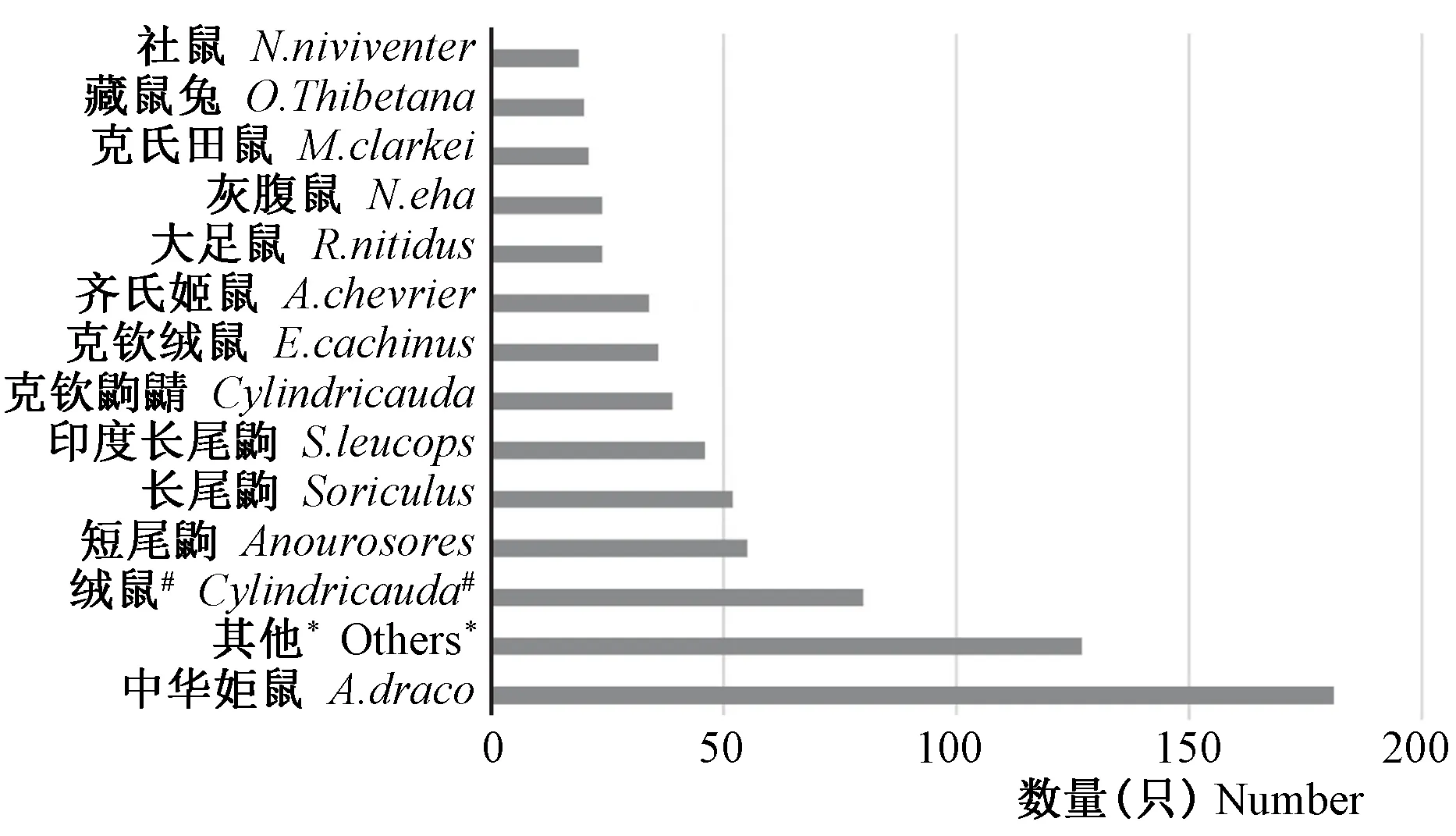
图1 捕获小型兽类的物种构成(其他*代表构成比低于2%的物种,包括中麝鼩、多齿鼩鼹、四川白腹鼠、高山鼩鼱、滇绒鼠、麝鼩、鼩鼱、褐腹长尾鼩、白腹鼠、大绒鼠、安氏白腹鼠、姬鼠、灰颈鼠兔、滇攀鼠、四川短尾鼩、鼠兔、白腹巨鼠、针尾鼹、大耳姬鼠、小麝鼩、田鼠、珀氏长吻松鼠;绒鼠#是指未明确其物种的绒鼠属的成员。)Fig.1 Species composition of small mammals captured in this study (Others* represents species with a composition ratio of less than 2%, including C. russula, N. gracills, N. excelsior, S. alpinus, E. eleusis, Crocidura, S. araneus, E. caudatus, R. coxingi, E. miletus, N. andersoni, Apodemus, O. forresti, Red Climbing Mouse, B. quadraticauda, Pikas, R.edwardsi, S. fusicaudus, A. lartronum, C. suaveolens, Microtus, D. pernyi; Eothenomys#represents unclear species in Eothenomys.)
2.2 病原体检测结果
分别使用无形体16S rRNA(rrs)基因通用引物、恙虫病东方体56 kDa基因、斑点热群立克次体17 kDa基因和柠檬酸合酶(gltA)基因及莱姆病螺旋体5S~23S rRNA间隔区基因的引物对541只小兽标本进行PCR扩增。当采用无形体rrs基因片段引物进行巢式PCR扩增时,共有17份标本出现目的条带,测序结果显示其中12份为新埃立克体CandidatusNeoehrlichiamikurensis、2份为沃尔巴克氏体Wolbachia、3份为巴尔通体Bartonella,阳性率分别为2.2%、3.7‰和5.5‰。12份新埃立克体分别来自7只姬鼠、3只绒鼠、1只鼩鼱和1只白腹鼠;沃尔巴克氏体来源于1只鼩鼱和1只灰腹鼠;3份巴尔通体分别来源于2只绒鼠和1只大足鼠。此外,用该引物扩增出的疑似阳性标本,在将PCR产物测序后发现,其中8份均为一种鼠类携带的肺炎支原体。采用恙虫病东方体56-kDa基因P10/P11引物进行PCR扩增时,发现4份标本(G243、G575、G728、G735)阳性,阳性率为7.4‰,分别来自白腹鼠、绒鼠、社鼠、社鼠。未发现斑点热群立克次体和莱姆病螺旋体阳性标本。
2.3 病原体基因特征分析
对部分rrs基因核苷酸序列Blast比对分析显示,检测到的12份新埃立克体,分别与分离自我国云南的Yunnan 06-50分离株、分离自日本的FIN686分离株以及分离自波兰的101 W分离株等的同源性为99.0%(Genbank号为GU227699、AB196304和MG670107);2份沃尔巴克氏体与wInc_Cu分离株(CP011148)同源性为98.0%;3份巴尔通体分别与分离自欧洲的AR 15-3、Houston-I和1-1C分离株(FN645485、CP020742和CP019489)同源性为99.0%。4份恙虫病东方体阳性标本之间核苷酸序列同源性为74.8%~97.5%。G728和G735核苷酸序列同源性为97.5%,与来自于韩国的Boryong血清型的2014-H30株同源性最高,均为97.0%。G243和来自于台湾的Kato-related血清型的GU446605同源性最高,为86.0%;G575与来自于韩国的Karp血清型2014-JN08同源性最高,为86.0%。进一步采用P34/P55引物对4份标本进行扩增,以获得更长的基因片段,结果从G243和G728 标本获得980 bp左右的基因片段。使用Mega 6.0运用邻接法绘制遗传进化树,如图2所示,G243和G735位于不同的分支,分别与Kato-related血清型的GU446605和Boryong血清型的L04956遗传进化关系最近。

图2 基于Orientia tsutsugamushi 56 kDa基因片段构建的进化树Fig.2 Phylogenetic tree based on O. tsutsugamushi 56 kDa gene fragment构建进化树所使用的软件为Mega 6.0,方法为邻接法,模型为Kimura-2-parameter model,Bootstrap值为1 000,图中黑色三角符号代表本研究中检测得到的序列。The NJ tree was constructed by Mega 6.0 program using the method of Kimura2-parameter model, and the bootstrap values was 1 000. The black triangle symbols in this figure represent sequences detected in the present study.
3 讨论
无形体科和立克次体科的病原体,如嗜吞噬细胞无形体、恙虫病东方体、斑点热群立克次体,为严格的细胞内寄生菌(Chapmanetal., 2006;Parolaetal., 2013),可由节肢动物叮咬脊椎动物后使其获得感染,并可致人患多种疾病(Chenetal., 1994;Dumleretal., 2005;Walkeretal., 2008)。据既往调查报道,云南省存在人粒细胞无形体病、恙虫病、Q热、流行性斑疹伤寒和斑点热等立克次体病,且为恙虫病和斑疹伤寒的高发区(Zhangetal., 2007)。
此次调查发现,贡山地区小型兽类中的优势属为姬鼠属和绒鼠属,这与既往在本地调查结果一致(龚正达等,1989)。从捕获的541只小型兽类中共发现4种病原体,这些病原体包括新埃立克体、恙虫病东方体、沃尔巴克氏体和巴尔通体。其中新埃立克体、恙虫病东方体、巴尔通体等3种病原体可由绒鼠携带,提示绒鼠作为当地优势鼠可携带多种致病性病原体,对当地自然疫源性疾病的传播有重要意义(Zhangetal., 2014)。
目前,无形体科主要包括无形体属、埃立克体属、新立克次体属、沃尔巴克氏体属、埃及小体属以及新埃立克体。本研究采用无形体科rrs基因通用引物进行扩增检测,除检测到12份新埃立克体、2份沃尔巴克氏体阳性外,还检测到3份巴尔通体。巴尔通体隶属于巴尔通体科(Bartonellaceae)巴尔通体属Bartonella,目前为人所熟知的巴尔通体所致疾病有猫抓病(Cat-scratch disease, CSD)、战壕热(trench fever)等(Bozhkovetal., 2014)。此次检测的巴尔通体是对无形体检测时,意外获得的结果,这与此前黎浩等(Lietal., 2012)在对恒河猴进行无形体检测时,意外检测到巴尔通体的结果一致。
12份新埃立克体阳性标本来源于多个种属,证实了该病原体在不同种属野生小型兽类间分布的广泛性。Blast比对和遗传进化分析显示,2份恙虫病东方体阳性标本属于Boryong血清型;1份与Karp血清型同源性最高,但仅为86.0%;另1份与Kato-related血清型菌株遗传进化关系最近,但同源性较低(86.0%),提示这两份标本可能为这两个血清型的变异株。此次调查进一步证实云南西北部地区小型兽类的物种多样性及其携带病原体的多样性,对相关自然疫源性疾病防控具有指导意义。
猜你喜欢
——贺《四川兽类志》正式出版
兽类学报(2023年6期)2023-03-11 19:43:25
中老年保健(2021年4期)2021-12-01 11:19:40
传染病信息(2021年6期)2021-02-12 01:52:12
自我保健(2019年5期)2019-08-02 03:54:24
中国人兽共患病学报(2016年10期)2016-01-31 01:26:14
西南医科大学学报(2015年1期)2015-08-22 13:02:00
中国当代医药(2015年9期)2015-03-01 02:02:09
——劳氏立克次体的研究现状
中国人兽共患病学报(2015年4期)2015-01-25 05:52:25
小学阅读指南·低年级版(2014年8期)2014-05-30 10:48:04
中国人兽共患病学报(2011年6期)2011-01-24 07:44:18
