
Effects of forest cover types and environmental factors on soil respiration dynamics in a coastal sand dune of subtropical China
2018-11-06 06:44WeiGaoZhiqunHuangGongfuYeXinjianYueZhiyongChen
Journal of Forestry Research 2018年6期
Wei Gao•Zhiqun Huang•Gongfu Ye•Xinjian Yue•Zhiyong Chen
Abstract Trees on sand dunes are more sensitive to environmental changes because sandy soils have extremely low water holding capacity and nutrient availability.We investigated the dynamics of soil respiration(Rs)for secondary natural Litsea forest and plantations of casuarina,pine,acacia and eucalyptus.Results show that significant diurnal variations of Rsoccurred in autumn for the eucalyptus species and in summer for the pine species,with higher mean soil respiration at night.However,significant seasonal variations of Rswere found in all five forest stands.Rschanged exponentially with soil temperatures at the 10-cm depth;the models explain 43.3–77.0%of Rs variations.Positive relationships between seasonal Rsand soil moisture varied with stands.The correlations were significant only in the secondary forest,and the eucalyptus and pine plantations.The temperature sensitivity parameter(Q10value)of Rsranged from 1.64 in casuarina plantation to 2.32 the in secondary forest;annual Rswas highest in the secondary forest and lowest in the pine plantation.The results indicate that soil temperatures and moisture are the primary environmental controls of soil respiration and mainly act through a direct influence on roots and microbial activity.Differences in root biomass,quality of litter,and soil properties(pH,total N,available P,and exchangeable Mg)were also significant factors.
Keywords Coastal sand dunes·Soil respiration·Secondary forest·Pine·Casuarina·Acacia·Eucalyptus plantations·Environmental factor·Q10
Introduction
Soil respiration is a major way in which CO2is released from terrestrial ecosystems to the atmosphere and has an important role in the global carbon cycle.The amount of carbon released globally by soil respiration accounts for about 60–90%of the total ecosystem respiration by terrestrial ecosystems(Schimel et al.2001).This is more than ten times the carbon released from fossil fuel combustion(Bond-Lamberty and Thomson 2010;Hashimoto 2012).Therefore,information about soil respiration and its environmental controls is of great significance for estimating future CO2concentrations in the atmosphere and its influence upon global climate change.
Total respiration from forest soils is from autotrophic respiration by roots and their associated rhizosphere micro flora,and by heterotrophic respiration generated by microbial consumption and the metabolism of soil organic matter.Soil respiration varies in different forest types because of the varying influence of tree species on biotic and abiotic factors(Fu et al.2013;Muñoz-Rojas et al.2016;Chen et al.2014;Song et al.2013;Wang et al.2013).However,the main causal factors for this varying influence are unclear.Different forest types can have different dominant factors such as soil temperatures and soil moisture.Differences in substrate availability(e.g.,organic carbon and nutrients)and soil carbon input(e.g., fine root biomass and turnover rate of litter)can result in temporal and spatial variation of soil respiration across forest types(Sheng et al.2010;Zeng et al.2014;Wang et al.2010;Raich and Tufekcioglu 2000).Therefore,more information on soil respiration is needed to understand the relationship between carbon cycling and specific forest types.
Sand dunes are common on the southeastern coast of China;the infertile soil has low water-holding capacity and high salinity.In addition,salt mist and frequent strong winds also limit forest growth and distribution(Chang et al.2011;Lin et al.2014).Casuarina equisetifolia L.is an actinorhizal nitrogen- fixing species with good wind and salt resistance and is often used for the establishment of protection forests in coastal sandy areas.The area of C.equisetifolia plantations in China is approximately 3×105ha(Zhong et al.2005)and play an important role in local environment improvement and ecological security maintenance(Xiao et al.2009).However,there are problems with adequate regeneration and productivity decline due to monoculture plantings and this has reduced their ecological benefits.To increase species diversity and improve the resistance of coastal protection forests,species from the Acacia,Eucalyptus,and Pinus genera have been introduced beginning in the 1960s.These are now established and are important components of coastal protection forests.In addition,a typical secondary forest is dominated by Litsea glutinosa(Lour.)C.B.Rob.,a native species with a patchy distribution in coastal sandy areas due to long-term protection following anthropogenic activities(deforestation or reforestation)and natural disturbances(e.g.,typhoon or sand burial).
The sandy coast is sensitive to environmental changes so different forest types may have different influences on ecosystem carbon cycling.This has been studied(Chen et al.2005;Lin et al.2014;Chang et al.2011)but information on soil respiration remains limited(Xiao et al.2009).This is especially true for environmental factors and variations in soil respiration across forest types which limit our ability to assess the role of coastal ecosystems in the global carbon balance.In this study,we investigated the variability of soil respiration under five typical forest covers,(a secondary forest of L.glutinosa,casuarinas,pine,acacia and eucalyptus plantations)in the southeastern coast of Fujian province.Our objectives were to:(1)describe the dynamics of soil respiration under different forest covers;(2)analyze the correlations of soil respiration with soil temperatures and soil moisture;(3)compare the Q10among the different forest types;and,(4)identify biotic and abiotic factors affecting soil respiration.
Materials and methods
Study site
The research was carried out at the Chishan Forestry Center of Dongshan County(23°38′N,117°24′E)in southeast Fujian Province.This region has a subtropical maritime monsoon climate,with annual mean temperatures ranging from 13.1 °C in January to 27.3 °C in July.Mean annual precipitation and evapotranspiration are 1104 and 2028 mm respectively.May to September is the rainy season when 61%of the rainfall occurs,and the dry season is usually November to February.The soil is a typic udipsamment with low nutrient content and thickness from 80–100 cm.
In December 2014, five forest covers including secondary forest,casuarina,pine,acacia,and eucalyptus plantations were selected as experimental plots.These stands had similar elevations and soil types,and average slopes were<10°.The main characteristics and topsoil properties of the 5 forest covers are listed in Table 1.
The secondary forest is native vegetation resulting from secondary succession with distinct tree,shrub,and herbous layers,which regenerates naturally and has been afforded long term protection by local people.The age of the secondary forest is more than 50 years,and the dominant species in the canopy are L.glutinosa and Celtis sinensis Willd.,the shrub layer is predominantly Glochidion eriocarpum,Litsea rotundifolia,Bridelia tomentosa,and Anodendron affine(Hook.et Arn.)Druce,and the main herb species are Ophiopogon bodinieri and Arthraxon hispidus var.hispidus.The casuarina and acacia plantations were established in 1992.The pine plantation was planted in 1993 with Pinus elliottii Engelm.or slash pine;the eucalyptus plantation was established in 2004 with Eucalyptus urophylla×E.grandis crosses,the previous species on this site was P.elliottii planted in 1976.All canopy densities of the different plantations were greater than 0.7,with few shrubs and grasses in the understory.
Experimental design and measurement
Four 20 m×20 m fixed plots were established in each of the forest stands in December 2014;each plot was separated by a 10 m buffer.Four PVC soil collars(inner diameter 20 cm and height 7 cm)were randomly inserted into the soil in each plot for soil respiration(Rs)measurement.The insertion depth was 4 cm at each sampling point.
Soil respiration(Rs)was measured using an automated soil CO2flux system(Li-8100,Li-Cor Inc.,Lincoln,NE,USA).The Rsrates between 09:00 and 11:00 am have been shown to be useful for estimating the daily mean CO2efflux rate(Xu and Qi 2001).Therefore monthly measurements were conducted between 09:00 and 11:00 am(Beijing time)on sunny days in every month from January 2015 to April 2016.In addition,diurnal measurements were taken at 2-h intervals from 9:00 am to 7:00 pm in April,July,and October 2015,and February 2016.Before sampling,the plants in the collars were carefully cut at ground level.
Soil temperatures at 10-cm depth near each collar were measured simultaneously with Rsusing a hand-held longstem thermometer(Model SK-250WP,Sato Keiryoki Mfg.Co.Ltd,Tokyo,Japan),and moisture in the 0–10-cm layer was measured using a time-domain reflectometry(TDR)probe(Model TDR300,Spectrum Technologies Inc.,Plain field,IL,USA).Final values represent the mean of three readings.
Five litter traps(0.5 m×0.5 m)with 1 mm nylon mesh were arranged randomly 0.3 m above the forest floor in each plot for litter collection.Beginning in March 2015,monthly litter was collected for determining annual litterfall.
In July 2015,ten soil cores(1.5 m length×5 cm diameter)were collected along the diagonal of each plot from the soil surface to the 100-cm depth.Fine roots(<2 mm diameter)were washed out and the remaining fine-root biomass was estimated using the method described by Yang et al.(2004).
Ten soil pits along the diagonal of each plot were used to collect soil in the top 10-cm depth in December 2015,and a 500 g sample from each plot was collected,and 300 g air-dried,ground,and sieved through a 2-mm sieve for microbial biomass,available P and exchangeable Mg analysis.The remaining soil was ground and sieved through a 0.149-mm sieve for soil organic carbon(SOC)and total N analysis.
Soil bulk density in the top 10-cm layer was determined using intact soil cores.Microbial biomass was analyzed by the chloroform fumigation-extraction method(Brookes et al.1985).The organic C content in the extract solution was measured using a TOC-VCPH/CPN Analyzer(Shimadzu,Japan)and converted to microbial biomass C(MBC)using a conversion factor of 0.45(Joergensen 1996;Wu et al.1990).The SOC and total N were determined using an elemental analyzer(Vario EL III,Elementar Analysensysteme GmbH,Hanau,Germany).Soil pH was measured with a pH meter.Available P was determined using Mo-Sb colorimetry and exchangeable Mg by atomic absorption spectrometry(Bao 2000).
Data analysis
Each plot was an experimental unit,thus replicate data were averaged by plots for analysis.One-way analysis of variance(ANOVA)with Duncan’s multiple comparison method were used to test the significance of differences in Rs,soil temperature,and soil moisture between different forest stands using a significance level of α=0.05.
In order to describe the relationship between seasonal Rs,soil temperature,and soil moisture,linear and nonlinear regression models were used as follows:

where T is soil temperature(°C)at 10-cm depth,W is soil moisture within the 0–10 cm depth(vol%),a,b,and c are undetermined parameters.
The temperature sensitivity parameter was calculated by the following:

where b is the parameter in Eq.(1)
The annual Rsin each plot was estimated by extrapolating measurements to monthly fluxes and summing from March 2015 to February 2016(Sheng et al.2010).Linear regression was used to estimate relationships between annual Rsand litter C,litter N,litter C:N ratios,SOC storage,total N,soil C:N ratios,pH,root biomass,MBC,available P and exchanged Mg.All statistical analyses were performed using the PASW Statistics 18.0 for Windows.
Results
Environmental variables
Mean monthly air temperatures during the research period showed a typical seasonal pattern with a peak in June.The highest monthly mean air temperature was 27.8°C and the lowest was 13.0°C;the mean annual air temperature was 20.3°C.Annual precipitation in 2015 was 1235 mm,with a rainy season from May to September when 77.4%of the total rainfall occurred.Monthly precipitation from January to April in 2016 was higher than in the same months in 2015,especially in April 2016 which had the second highest rainfall(July 2015 was highest)(Fig.1).

Fig.1 Monthly cumulative precipitation and monthly mean air temperature during 2015–2016
There was no significant difference in litterfall among the five forest types.The litter C content was highest in the acacia plantation and lowest in the secondary forest;litter N content was highest in secondary forest,followed by the acacia plantation,and lowest in the pine plantation.Litter C:N was highest under pine and lowest in the secondary forest and the acacia plantation.Fine root biomass was highest with casuarina and in the secondary forest and lowest with pine.Soil pH was highest in the secondary forest followed by the eucalyptus plantation.Soil SOC storage,total N,available P,exchangeable Mg and MBC were all highest in the secondary forest;no significant differences were found among the other forest stands.Soil C:N ratio was highest in the eucalyptus planation followed by the casuarina plantation,and lowest under acacia(Table 1).
Diurnal variation of soil respiration
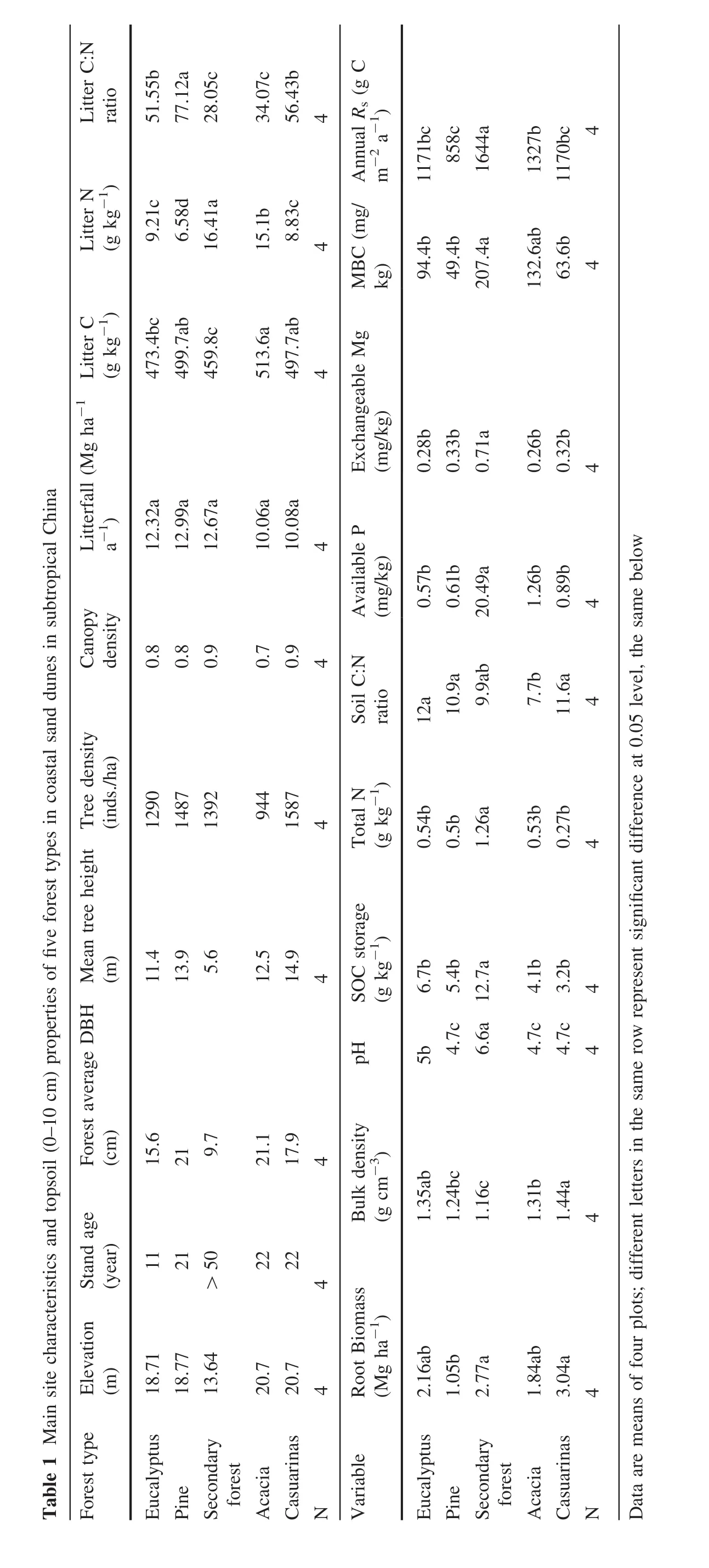
Significant diurnal variation in Rswas observed with eucalyptus during autumn and with pine during the summer(P<0.05).Daily variation had a minimum value at 15:00 h and a maximum between 23:00 and 1:00 h.No significant diurnal variation in Rswas observed in the secondary forest,or in the acacia or casuarina plantations(P>0.05).Most of the mean soil respiration rates at night were higher than during the daytime(Fig.2),yet these were inconsistent among the periods measured.
Seasonal variations of soil respiration,temperature soil moisture
Seasonal variations of soil temperature at the 10-cm depth were highest in the summer and lowest in the winter months.Higher temperatures from May to September gradually decreased,with the lowest values occurring the following January.Seasonal coefficients of variation(CV)in soil temperature were 23.1,22.1,25.2,26.3 and 24.1%for plantations of eucalyptus,pine,acacia,and casuarina and for the secondary forest,respectively.The annual mean soil temperatures under the different plantations and secondary forest ranged from 21.9 to 22.2°C with no significant differences between cover types(P>0.05).
The highest soil moisture readings were observed in the transition period from spring to summer(April or May),and the lowest occurred from July to November.Seasonal CV values were 56.7,36.3,40.0,35.2,and 37.2%for plantations of eucalyptus,pine,acacia,and casuarina and for the secondary forest,respectively.Annual mean soil moisture ranged from 3.3 to 4.4%(Fig.3,means not shown);no significant differences were found between cover types(P>0.05).
The monthly Rsshowed seasonal variabilities,peaking in eucalyptus and acacia plantations and in the secondary forest in June,while the highest values in the pine and casuarina plantations occurred in September.All values were lowest in December or January,2015.A sharp increase in Rsoccurred in April 2016,which might have been due to the abundant rainfall(monthly total precipitation was 282 mm)along with rising spring temperatures.The annual mean Rsin the secondary forest was significantly higher than in plantations.There was no significant difference in annual mean Rsamong acacia,eucalyptus or casuarina plantations.Seasonal CV of Rswas 44.5,42.0,39.7,33.6 and 67.1%for plantations of eucalyptus,pine,acacia and casuarina and for the secondary forest,respectively.
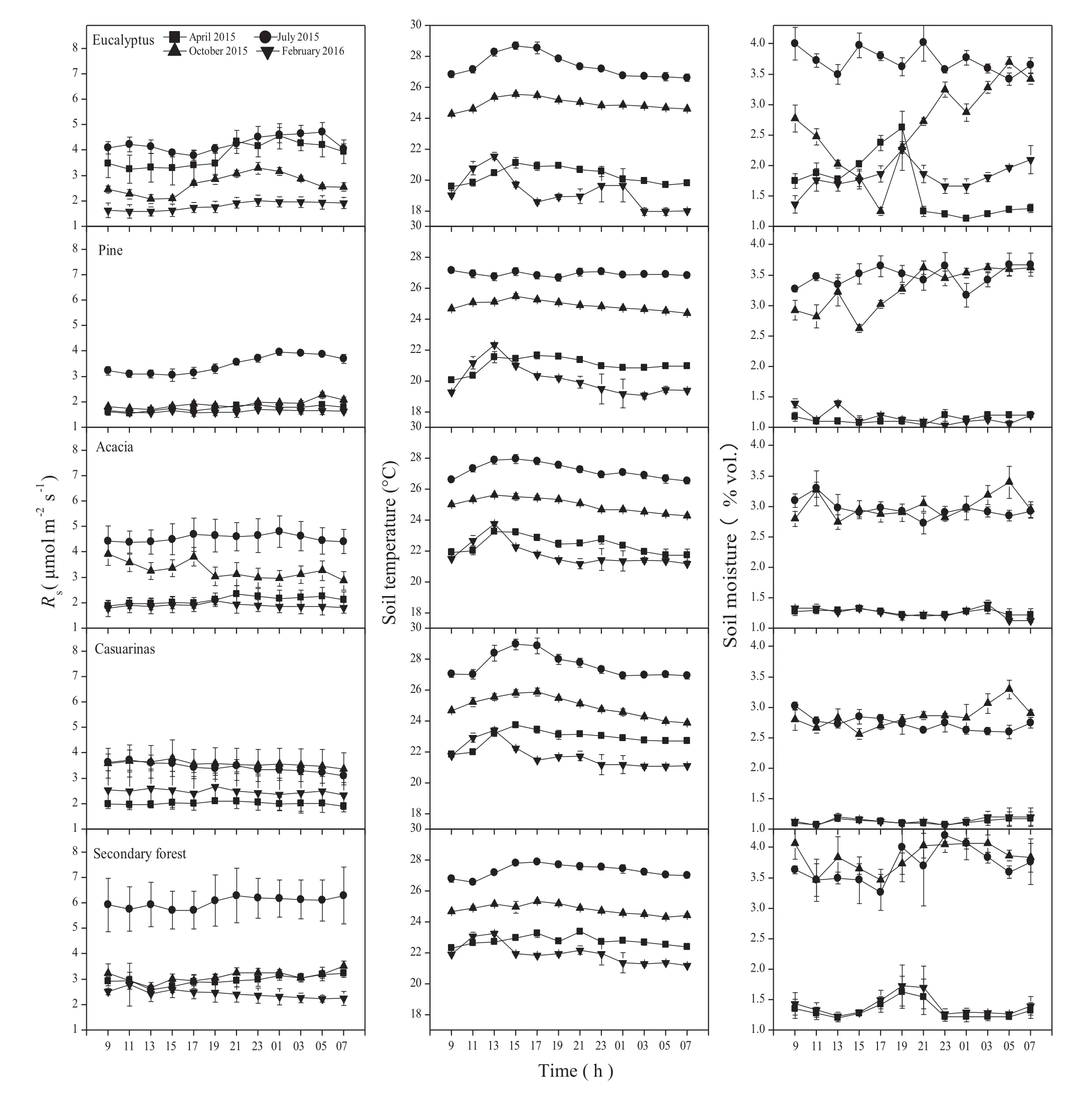
Fig.2 Diurnal variation of soil respiration,soil temperature and soil moisture for five forest types in April 2015,July 2015,October 2015 and February 2016,vertical bars represent standard error
Effects of soil temperature and moisture on soil respiration
The monthly Rshad a positive exponential correlation with T10for all sites(Fig.4),which explained 43.3–77.0%of the temporal variation of Rsfor the different cover types.The highest R2coefficient was in the pine plantation and the lowest with acacia.The Q10value was highest in the secondary forest and the lowest in the casuarina plantation(Table 2).
Correlations of monthly Rswith W10were weaker than those with T10.A significant linear correlation was found for the secondary forest,and for plantations of eucalyptus and pine(Fig.4),accounting for 15.0–22.9%of the temporal variation of Rs(Table 2).This indicates that the temporal variations of Rsin these three cover types were significantly controlled by soil moisture.There was no obvious relationship between Rsand W10in plantations of acacia and casuarina.The bi-factor model that integrates W10with T10improved R2compared to the T10-based model,which could explain 48.4–78.1%of the temporal variation of Rsfor different cover types(Table 2).
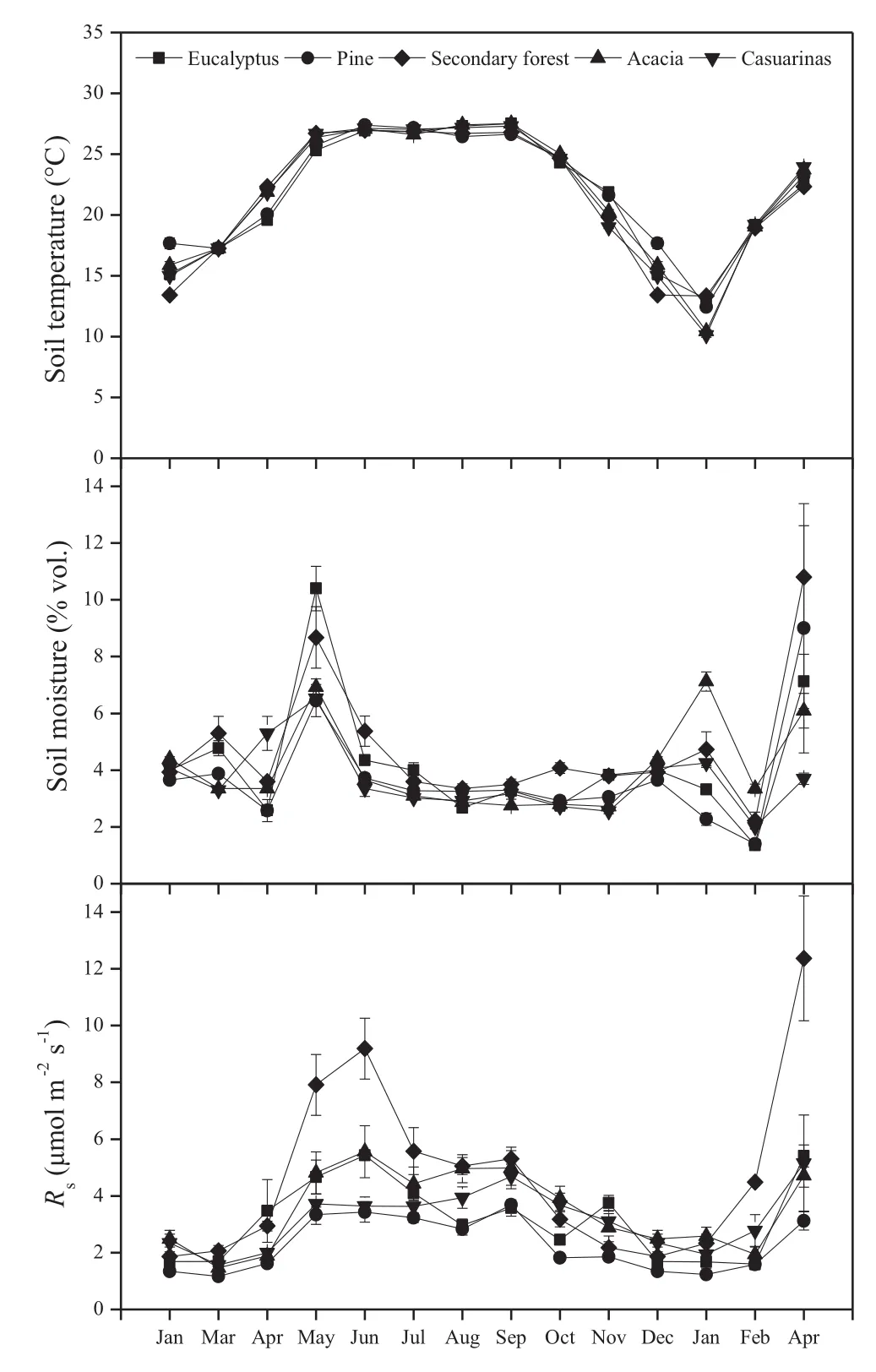
Fig.3 Monthly soil temperature(°C)at 10 cm depth,soil moisture(vol%)in the 0-10 cm layer and soil respiration(μmol m-2s-1)for five forest types during 2015–2016
Relationship between annual soil respiration and biotic and abiotic factors
Annual Rswas 1171±63.4,858±33.9,1327±151.86,1170±68.7,and 1644±143.5 g C m-2a-1for plantations of eucalyptus,pine,acacia,and casuarina,and for the secondary forest,respectively,and was significantly higher in the secondary forest than in the plantations.The annual Rsin different plantations were highest with acacia and lowest with pine,with a significant difference between the two sites.No significant differences were found between other plantations(Fig.5).The mean annual Rshad a significant positive correlation with litter N and MBC,but a significant negative correlation between Rsand litter C:N among different cover types(P=0.013,Table 3).No significant correlation of mean annual Rswith other biotic and abiotic factors was found.
Discussion
Diurnal and seasonal dynamics of soil respiration
Little diurnal fluctuation in soil temperatures was found in the five cover types of this coastal sandy area because of the closed-tree canopies,so diurnal variation of Rsin this area might be less sensitive to soil temperatures.Significant diurnal variation in Rswas only observed in the autumn for the eucalyptus plantation and in summer for the pine plantation.However,soils in this sandy area lack sufficient water-holding capacity,and mean annual soil moisture only ranged from 3.3–4.4%(Table 2),indicating that soil water deficit may be an important limiting factor for soil respiration.In this coastal sandy area,higher relative humidity at night might favor microbial activity,leading to increased levels of Rs.
Seasonal variation was recorded in different cover types and is consistent with studies in different ecosystems(Chen et al.2014;Sheng et al.2010;Liu et al.2011;Zhang et al.2013;Zeng et al.2014).Seasonal variations in Rsare apparently controlled by soil temperatures and moisture at the ecosystem level.
Correlation of seasonal soil respiration with soil temperatures and moisture
Soil temperatures can influence seasonal Rsby affecting microbial activity,root growth and decomposition of litter(Chen et al.2010a,b;Bond-Lamberty and Thomson 2010;Han et al.2016).We found a significant exponential relationship between seasonal Rsand soil temperatures,indicating that Rscould be estimated by soil temperatures alone.This is consistent with previous studies(Sheng et al.2010;Raich and Schlesinger 1992;Song et al.2013).The temporal variation of Rscould be explained by soil temperatures from 43.3%in the acacia plantation to 77.0%in the pine.The difference in temperature controls may be caused by the quality of substrate,root growth,and microbial activity(Chen et al.2010a,b).
Fig.4 Relationships between soil respiration(Rs)and soil temperature(T10)at 10 cm depth and soil moisture(W10)in the 0–10 cm layer for five forest types in coastal sand dunes in subtropical China
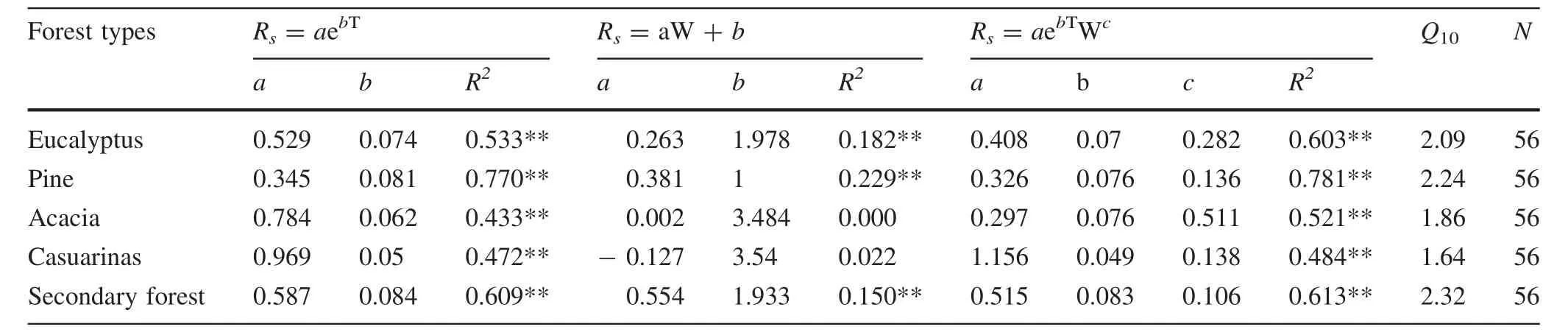
Table 2 Regression models of monthly soil respiration(Rs)against soil temperature(T10)and soil moisture(W10)and Q10values
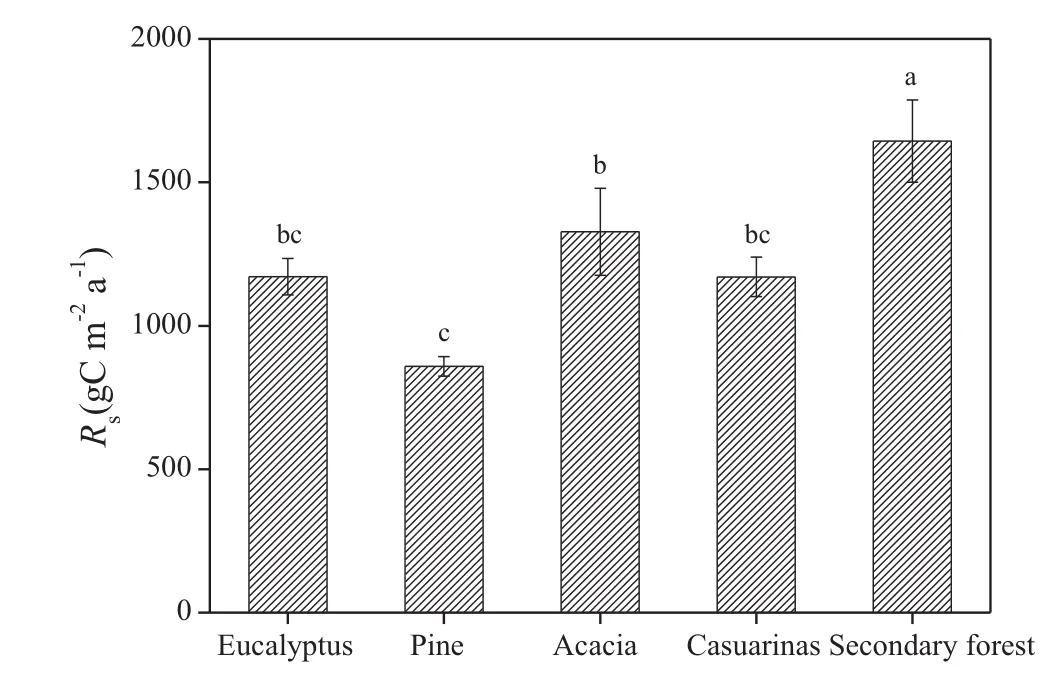
Fig.5 Annual soil respiration(Rs)for different forest types
The Q10value of Rsranged from 1.6 to 2.3 across the five cover types with a mean of 2.0 that was lower than the global average of 2.4(Raich and Schlesinger 1992).This might be due to limited microbial activities,root respiration and diffusion of soluble organic matter by lower soil moisture which made root and microbial respiration less sensitive to soil temperature variations.Similar results have been found in other studies(Chen et al.2010a,b;Zeng et al.2014).Lower Q10in the coastal sandy area indicates lower CO2emissions under continued global-warming compared to regions with higher Q10.Q10was highest in under secondary forest,indicating that Rswas more sensitive to soil temperatures.Previous studies suggested that the Q10is a combined response of soil temperature,soil moisture,SOC concentration,and ecosystem types(Zeng et al.2014;Chen et al.2014).Zheng et al.(2009)found that ecosystems with higher SOC generally had higher Q10,which was due partially because higher SOC supplied larger amounts of substrates for soil respiration(Talmon et al.2010).We found no significant differences in soil temperatures and soil moisture between different cover types;the high Q10in the secondary forest may have been caused by its higher SOC concentration(Table 1).
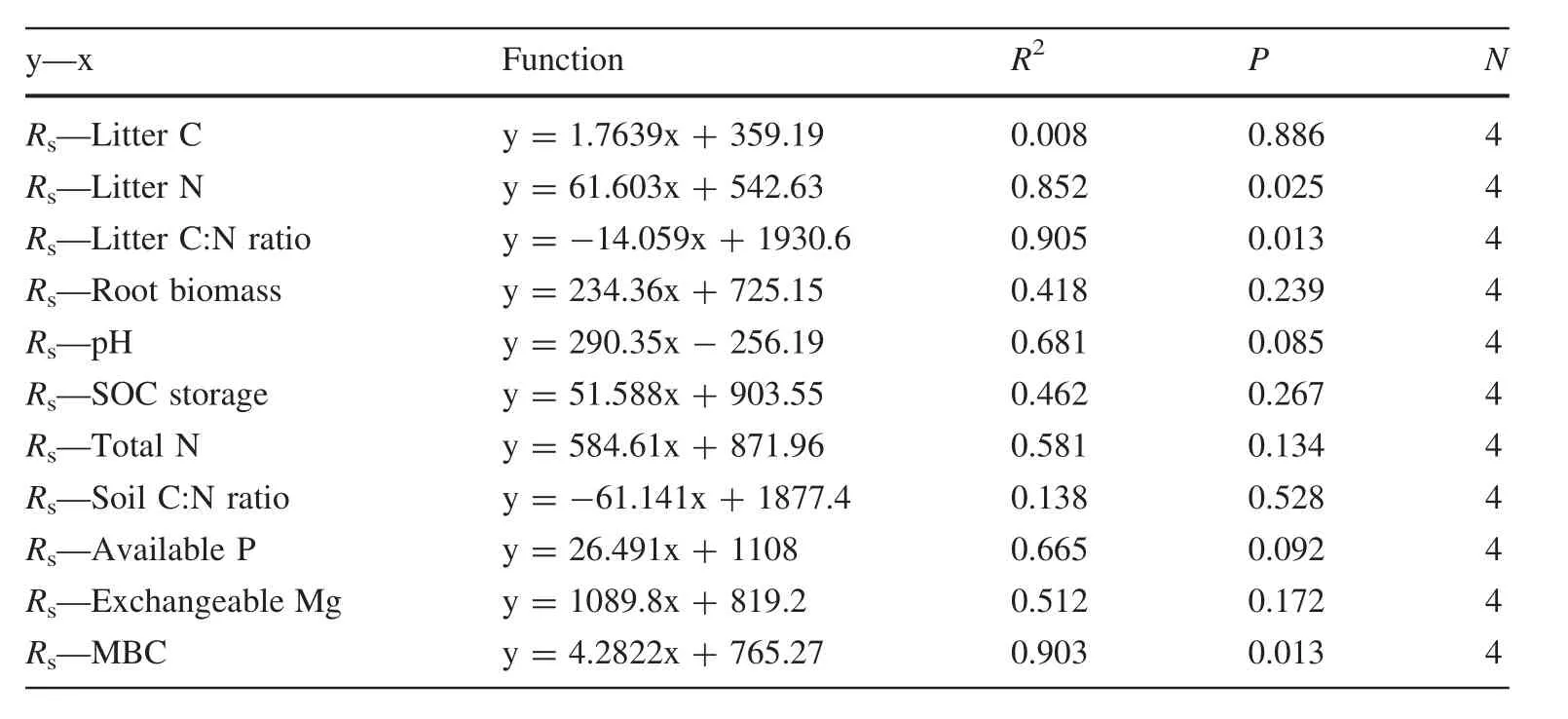
Table 3 Relationship between annual soil respiration(Rs)and biotic and abiotic factors
The relationship between Rsand soil moisture was complex compared to that with soil temperature.The correlation was significantly positive under the secondary forest,and in the eucalyptus and pine plantations,but not significant under acacia or casuarina,indicating that there were different relationships between Rsand soil moisture in different vegetation types.Similar results have been reported in different land-use types in mid-subtropical China(Sheng et al.2010)and in fire-affected semi-arid ecosystems in western Australian(Muñoz-Rojas et al.2016).The bi-factor models integrating W10with T10improved R2at different levels compared to the model based on T10only,indicating that soil temperature and soil moisture played an interactive or complementary role in influencing Rsin the coastal area,a phenomenon also seen in other regions(Zhang et al.2010;Sheng et al.2010).Although seasonal Rsvalues were mainly influenced by soil temperatures,soil moisture also exerted a strong effect.For example,high air temperatures from July 2015 to September 2015 may have led to high evapotranspiration rates and a reduction of soil water content which limited the response of roots and microbial respiration to soil temperature.Rsgenerally peaked in the periods with both high soil temperatures and high soil moisture(May 2015,June 2015 and April 2016).The results presented support previous studies showing that low soil moisture constrains soil CO2ef flux(Xu and Qi 2001).
Rainfall can cause pulse effects on soil respiration because the number and activity of soil microbes generally increase after a suitable rainfall(Xu and Qi 2001;Lee et al.2002;Lee and Wen 2006).The monthly precipitation in April 2016 was 282.2 mm(Fig.1)which was second only to the amount in July 2015.Besides increased soil temperatures in spring,soil moisture increased because of abundant rainfall,and Rsin the different forest covers increased accordingly in a small pulse during April 2016.
Effect of forest cover types on annual soil respiration
The annual Rsranged from 858 to 1644 g C m2a-1under the different cover types.It was highest in the secondary forest,lowest in the pine plantation,and averaged 1234 g C m-2a-1in the coastal area.Annual Rsvalues for plantations of eucalyptus,acacia,and casuarina and for the secondary forest were 26.7,35.3,26.7 and 47.8%higher,respectively,than the Rsfor the pine plantation,suggesting that the rates of soil respiration beneath broad-leaved forests are higher than those beneath conifers at the same sites.These results are consistent with previous findings(Huang et al.2014;Sheng et al.2010;Song et al.2013;Raich and Tufekcioglu 2000).The annual Rsfor the 22-year-old casuarina plantation was 1107 g C m-2a-1;this value is within the range of the annual Rsfor 6-to 31-year-old casuarina plantations reported by Xiao et al.(2009).
The root biomass under pine was lower than for other forest covers(Table 1).This may indicate lower root respiration and explain the lowest annual Rsunder pine.Soil respiration and soil organic carbon accumulation were also influenced by the quantity and quality of litterfall(Sheng et al.2010).We found no significant difference in annual litterfall between the different cover types.However,differences in litter N and C:N ratios were significant.The litter C:N ratio for pine,eucalyptus and casuarina plantations was significantly higher than that for the secondary forest and the acacia plantation(Table 1).The annual Rswas significantly positively correlated with litter N and negatively with the litter C:N ratio(Table 3).This indicates that soil respiration was significantly influenced by litter quality.In addition,litter in coniferous forests decomposes more slowly than litter in broad-leaved forests(Zhang et al.2008),and high tannin concentrations in C.equisetifolia branchlets could further reduce decomposition rates(Ye et al.2012).It is evident that substrate availability in pine and casuarina plantations was limited by the litter decomposition rate.Thus differences in root biomass,as well as quality and turnover rate of litter,might be causal factors(in addition to soil temperatures and moisture)affecting the annual Rsfor different cover types in the coastal sandy area.
Soil properties can be significantly influenced by vegetation type(Fu et al.2010;Lu et al.2014),and the composition of soil microbial communities by soil properties(Griffiths et al.2011;Fierer 2006;Lauber et al.2009;Uffen et al.2010;Landesman et al.2014),such as soil pH(Shen et al.2013;Feng et al.2014;Landesman et al.2014),total N(Tewary et al.1982),available P(Gallardo and Schlesinger 1994)and exchangeable Mg(Xu and Qi 2001).We observed higher soil pH under secondary forest and in the eucalyptus plantation.The total N,available P,and exchangeable Mg in the secondary forest were all significantly higher than in the other forest types,which might have caused a higher MBC in the secondary forest and indicate greater microbial activity than in the other cover types(Table 1).There was a significant positive correlation between annual Rsand MBC in the coastal area(Table 3).This data suggest that the influence of soil properties on microbial biomass and microbial activity might also affect annual Rs.
Conclusions
The dynamics of soil respiration Rswere investigated in five typical forest cover types(a secondary Litsea forest,and plantations of C.equisetifolia,P.elliottii,A.crassicarpa,and E.urophylla×E.grandis).Significant diurnal variations in Rswere observed in autumn for the eucalyptus plantation and in summer for the pine,with higher mean soil respiration at night.No significant diurnal patterns were found in other seasons in the other different cover types.
Similar seasonal variations of Rswere found in all five cover types and peaked in June to September and were lowest in December to January.The mean annual Rswas highest in the secondary forest and lowest in the pine plantation.
Seasonal variations of soil respiration were exponentially correlated with soil temperature at the 10-cm depth,and the relationship between seasonal Rsand soil moisture varied with tree species.The two-factor model integrating W10with T10improved R2at different levels compared to the model based on T10only,indicating that the soil temperature and soil moisture had interactive or complementary roles influencing Rsin the coastal area.The highest Q10in the secondary forest indicated greater sensitivity of Rsto soil temperature.
Annual Rswas highest in the secondary forest and lowest in the pine plantation.Our findings indicate that soil temperature and moisture exert primary environmental effects on Rs,mainly through direct influences on root and microbial activity.Root biomass,quality of litter(litter N and C:N),and soil properties including pH,total N,available P,and exchangeable Mg are also significant factors influencing different respiration rates.
 Journal of Forestry Research2018年6期
Journal of Forestry Research2018年6期
- Journal of Forestry Research的其它文章
- Black locust(Robinia pseudoacacia L.)as a multi-purpose tree species in Hungary and Romania:a review
- The impact of the environmental factors on the photosynthetic activity of common pine(Pinus sylvestris)in spring and in autumn in the region of Eastern Siberia
- Osmoregulators in Hymenaea courbaril and Hymenaea stigonocarpa under water stress and rehydration
- Effect of nitrogen levels on photosynthetic parameters,morphological and chemical characters of saplings and trees in a temperate forest
- Free amino acid content in trunk,branches and branchlets of Araucaria angustifolia(Araucariaceae)
- Exogenous application of succinic acid enhances tolerance of Larix olgensis seedling to lead stress