
Effect of Vachellia tortilis on understory vegetation,herbaceous biomass and soil nutrients along a grazing gradient in a semi-arid African savanna
2018-11-06 06:44TemesgenYadetaElmarVeenendaalKarleSykoraZewduTessemaAddisuAsefa
Journal of Forestry Research 2018年6期
Temesgen Yadeta•Elmar Veenendaal•Karle Sykora•Zewdu K.Tessema•Addisu Asefa
Abstract The spatial pattern and abundance of herbaceous vegetation in semi-arid savannas are dictated by a complex and dynamic interaction between trees and grasses.Scattered trees alter the composition and spatial distribution of herbaceous vegetation under their canopies.Therefore,we studied the effect of Vachellia tortilis on herbaceous vegetation composition,biomass and basal area,and soil nutrients on sites with varying grazing intensities in the central rift valley of Ethiopia.Data were collected on species composition,cover and biomass of herbs and grasses,and soil moisture and nutrient contents under light,medium,and heavy grazing pressures,both under the inside and outside of V.tortilis canopies.Species richness was similar in both locations but decreased with increased grazing.Only the overall biomass and herb cover were significantly greater under the canopy than outside,and overall biomass showed significant unchanging decline with increased grazing.However,vegetation cover was significantly greater on moderately grazed sites compared to low and heavily grazed sites.All soil variables were significantly higher under V.tortilis canopies than outside.Our findings suggest that V.tortilis has more effect on composition and diversity of herbaceous vegetation than on species richness,and that V.tortilis promotes the herbaceous layer biomass by reducing soil moisture loss and increasing soil fertility under the inside than outside the canopies.Therefore,we suggest that management practices should be directed on reducing pressure on V.tortilis by regulating grazing.Low to moderate grazing levels(i.e.,a stocking rate less than 39.6 TLU ha-1yr-1)seems to be tolerable to ensure sustainable conservation of the species in the study area in particular and in semi-arid savannas in general.
Keywords Basal area cover·Grazing pressure·Inside/outside canopy·Soil moisture
Introduction
The vegetation of semi-arid African savannas is characterized by a dominance of grasses mixed with scattered trees(Sankaran et al.2004).Accordingly,the spatial patterns and abundance of herbaceous vegetation are dictated by dynamic and complex interactions between trees and grasses(Scholes and Archer 1997).Scattered trees alter the composition and spatial distribution of herbaceous vegetation under their canopies(Belsky et al.1993a;Belsky 1994;Moyo and Campbell 1998).
Some studies suggest that tree canopies have a positive effect on understory vegetation by improving soil nutrient conditions(Belsky et al.1989;Jake and Michael 1990),reducing heat stress(Belsky et al.1989;Gebrekirstos et al.2008),and improving soil moisture content(Ludwig et al.2003).In contrast,other studies have reported negative effects and attribute their findings to the fact that trees compete with understory vegetation for water,light and nutrients(Ludwig et al.2001,2004a,b).Some of the reasons for such discrepancies among various studies might be due to the variation in the type of confounding factors such as livestock grazing(Abule et al.2005;Angassa and Oba 2010;Tessema et al.2011;Tessema and Belay 2017).Several studies suggest that regulated and moderate levels of grazing often lead to increased understory vegetation(Dyer 2002;Treydte et al.2007;Angassa and Oba 2010;Hailu et al.2015).However,studies aiming to promote better understanding of the relationships between tree canopies and understory vegetation should be carried out by taking into account the effects of potential confounding variables,including grazing intensities.Studies undertaken,particularly at a local level,are of paramount importance as effective conservation and management decisions for dominant species are often made at local levels.Due to the significant biodiversity conservation and ecosystem services of dominant trees,coupled with the ever-increasing environmental degradation in semi-arid ecosystems,studies that contribute to the sustainable management of these ecosystems and to the socio-economic development of the local people are of paramount importance.However,land and vegetation degradation have been increasing at an alarming rate due to continuous heavy grazing,deforestation for crop cultivation,charcoal making,fuel wood consumption and construction materials(Senbeta and Tefera 2001;Treydte et al.2007).
Vachellia tortilis(Forssk.)Galasso&Ban fi(umbrella thorn acacia)is one of the most dominant tree species in the rift valley of Ethiopia,typical of semi-arid African savannas.The species is increasingly threatened by overharvesting for charcoal making,fuel wood consumption and construction materials(Eshete and Sathi 1998;Gebrekirstos et al.2008;Mengesha et al.2011;Tessema et al.2011),V.tortilis would be an interesting study species from ecological and conservation perspectives(Senbeta and Tefera 2001;Abule et al.2005;Tessema and Belay 2017).The species has been recommended for agroforestry as it provides direct and indirect ecosystem services to local communities(Eshete and Sathi 1998),improve aggregate soil particles(Mengesha et al.2011),and promotes the growth of herbaceous vegetation for domestic and wild herbivore foraging in semi-arid environments(Abule et al.2005;Tessema et al.2011).
However,the information about the effects of V.tortilis on understory plant communities and soil parameters in relation to the effects of various grazing intensities is minimal.Therefore,we studied herbaceous vegetation composition,cover,biomass,and soil nutrients along grazing gradients(i.e.,light,medium,and heavy),under the inside and outside canopies of mature V.tortilis.We hypothesize that the contribution of V.tortilis canopies on understory vegetation,basal area and biomass is more pronounced under intermediate grazing than under low or heavy grazing.We compared soil moisture and nutrient contents(phosphorus,carbon and nitrogen)between canopy and grazing treatments,and evaluated their relationships with vegetation cover and herbaceous biomass.
Materials and methods
Study areas
The study was conducted in Abijata Shalla Lakes National Park(7°31′N,38°39′E)and the nearby Abernosa Cattle Breeding Ranch(7°46′N,38°38′E),both situated in the central rift valley of Ethiopia,typical of semi-arid environments.Abijata Shalla Lakes National Park is situated approximately 200 km south of Addis Ababa,the capital of Ethiopia.The park was established in 1970 with the aim of conserving high diversity of bird species inhabiting the area.The total area is 887 km2,over half(482 km2)is covered by Abijata Lake and Shalla Lake(Mengesha et al.2011;Kumssa and Bekele 2014).The Abernosa Cattle Breeding Ranch was established 42 years ago to improve the productivity of local cattle through crossbreeding with improved exotic breeds of Holstein–Friesian and Jersey cattle(Tessema et al.2011,2012).The current extent of the Ranch is~26 km2(pers.comm.with the ranch administration).Both the Park and Ranch areas are at similar altitudes(1500–1657 m a.s.l.)and have similar dominant plant species(Argaw et al.1999).The Ethiopian central rift valley is characterized by a semi-arid savanna ecosystem where several Vachellia species occur,such as V.tortilis,V.senegal(L.)Britton(gum acacia)and V.seyal(Dellie)P.J.H.Hurter(red acacia),and other woody species such as Dichrostachys cinerea Wight et Arn.(Sickle bush)and Balanites aegyptiaca(L.)Delile(desert date)(Kumssa and Bekele 2014).The peak rainfall season is from June–September while the dry season lasts from October–February.The mean annual rainfall is estimated to be 650 mm(Mengesha et al.2011).
Similar to other protected areas in Ethiopia,the Abijata Shalla Lakes National Park has been under severe pressures from human settlement,crop cultivation and livestock grazing.Of the total area of 405 km2,only 31 km2(~8%)is free of cultivation and settlement(EWNHS 2009).On average,191,982 heads of livestock(132,629 cattle,40,490 sheep and goats and 10,153 horses)use the Park every day(ASLNP,2010 unpubl.data).However,in a 1.2 km2area of the Park,wire-fenced since its establishment,the density of herbivore wildlife species is relatively low.This area is better protected from livestock grazing and logging activities that occur sporadically(Kumssa and Bekele 2014).Only three wild herbivores use the fenced area;the common warthog(Phacochoerus africanus Gmelin.),Grant’s gazelle(Nanger granti Brooke)and Reedbuck(Redunca arundinum Boddaert),with estimated populations of 32,58 and 3 ind.,respectively(ASLNP 2010;unpubl.data).For the purpose of this study,we selected the wire-fenced portion of the Park as representative of light grazing(i.e.,no domestic animal gazing and only a few wild herbivores)and the unfenced part(i.e.,the 31 km2area free of cultivation and settlement)as representing a heavy grazing site(465.2 TLU ha-1yr-1:TLU=tropical livestock unit is an animal weighing 250 kg).In contrast,the Ranch was selected as a moderately grazing site as it supports a lower number of livestock than the site designated as under heavy grazing,but a greater number than the site with light grazing since it currently supports approximately 39.6 TLU ha-1yr-1(Yadeta T,pers.comm.with the Ranch administration).
Data collection
Individual tree selection
An individual V.tortilis tree was measured for diameter at breast height(DBH),and based on the nearest-neighbor method,the DBH of 20 ind.trees in each of the three grazing treatment sites were measured.Trees were selected using two suitability criteria:size and isolation.A given individual was considered suitable if it had a DBH>10 cm(Newton 2007),and was isolated by>50 m from other trees and/or dense shrubs.Accordingly,for the three sites,mean(±SD)tree heights were 8.3 m(0.8),8.4 m(0.5)and 9.8 m(0.6),and DBH 30.5 cm(10.4),38.3 cm(8.0)and 36.9 cm(10.0).Crown areas(m3)were 25.5(1.5),25.5(1.4)and 26.4(1.0),respectively for the low,heavy and moderate grazing sites.
Sampling of herbaceous vegetation
Data were collected between September and November 2010.For each individual tree,one transect line originating from the stem to open land(i.e.,depending on the radius of the under canopy),was randomly positioned in any of the four cardinal directions.Vegetation data were sampled in four 1 m×1 m quadrats established at 25 and 75%(representing areas under the canopy)and at 125 and 175%(representing areas outside canopy)relative to canopy radius(canopy radius=100%)(Fig.1).Attempts were made to avoid nearby dense shrubs and termite mounds when transects were positioned.Cover was visually estimated for each species and for each plant life-form(i.e.,grasses and herbs).Species identification was made in the field and by comparing herbarium materials in the Park education centre.Vegetation nomenclature followed Phillips(1995)and Kelecha(1987).Kyalangalilwa et al.(2013)was used for the recently revised genus Acacia Miller s.l.(Fabaceae:Mimosoideae).
To determine aboveground biomass,vegetation in each quadrat was clipped to the ground at stubble height,excluding stem base,corms,and/or roots.The data were collected at the end of the rainy season,a period when vegetation production peaks,i.e.,most grasses were in flowering and seed setting stages.The samples were separated into grasses and herbs.Fresh weights were taken using a handheld 1000-g capacity field balance immediately after clipping and sorting.These samples were airdried and weighted to determine dry biomass.The averages of quadrat one and two(25 and 75%,under canopy)and of quadrat three and four(125 and 175%,outside canopy)were calculated for each individual tree and used for comparisons.
Soil sampling and analyses

Fig.1 Sample design for data collection under and outside tree canopies;species composition and vegetation biomass data taken in 1 m×1 m plots laid out along a transect line originating from the tree trunk to open land at 25 and 75%(representing areas under crown)and at 125 and 175%(representing areas outside crown)relative to crown radius(crown radius(r)=100%)
Soil samples were taken in the quadrats established at 25%(under canopy)and 175%(outside canopy)along each transect where the vegetation data were collected(Fig.1).The samples were bulked into one composite sample under the inside and outside canopies in each grazing gradient,weighted immediately after collection and dried at 105°C to constant weight.Samples were analyzed for moisture content(gravimetrical by drying at 105°C),total phosphorus(digesting in tubes with H2SO4-salicylic acid-H2O2and selenium),total nitrogen,and total carbon(Fisons CNH-S analyser)(Jones et al.2006).
Data analyses
Species composition
Prior to species richness comparisons,rarefaction curves were used to examine sampling completeness in each site of the two main treatment categories,as well their interactions(e.g.,light grazing under canopy),using the Estimate S v.9.1.0 software(Colwell 2013).Sampling is adequate if the rarefaction curves reach an asymptote or if they closely converge with a richness estimator(Colwell 2013).We used Chao2 richness estimator,the most reliable for presence/absence data(Colwell 2013)to estimate the expected number of species that would be found in each site.
Multivariate community analysis was implemented in PRIMER software(Clarke and Gorley 2006)to assess differences in species composition of the herbaceous vegetation between canopies and grazing level categories.The similarity in composition of the herbaceous vegetation was calculated using Sorensen’s similarity index using species presence/absence data(Clarke and Gorley 2006).This is the most useful for ecological community data(McCune and Mefford 1999).Using this similarity matrix(i.e.,similarity values between each possible pair of sample combinations)as input,we then used a two-way crossed analysis of similarity(ANOSIM)to examine the effect of V.tortilis canopy and grazing pressure on the composition of the herbaceous layer.Similarly,we used one-way ANOSIM to examine the effect of canopy on species composition in each grazing level.Global R-values were used to determine the degree of similarity among treatments.This is a non-parametric permutation procedure applied to rank similarity matrices underlying sample ordinations(Clarke and Gorley 2006).The closer the value of R is to 1,the more dissimilar species assemblages are.Furthermore,to identify species that characterize each treatment,similarity percentage analysis(SIMPER)was conducted in PRIMER software to calculate the percentage contribution each species made to the dissimilarities between each possible pair of assemblages(e.g.,between light-grazed and heavy-grazed sites)(Clarke and Gorley 2006).This was based on percent ground cover values of each species at each sampling point.
Herbaceous biomass and basal area cover
We used the Generalized Linear Model with identity link function and normal distribution to examine the effects of canopy and grazing pressure on aboveground biomass and basal area of overall herbaceous vegetation,(i.e.,by pooling herbs and grasses together and separately).Proportional data(vegetative cover)were arcsine transformed to meet the assumption of normality and homogeneity of variances prior to carrying out the analysis.Mean differences between grazing levels were tested using the Bonferroni adjustment for multiple comparisons(Quinn and Keough 2002).The interaction terms fitted in each model showed non-significant results in all cases.Thus,we report and discuss our results in the context of main effects.Similar procedures were followed to test the effects of canopy and grazing and their interaction on soil moisture and nutrient contents.Finally,we used a Multiple Linear Regression Model to assess the relationships between each vegetation variable and all soil variables.All these analyses were carried out in SPSS version 20 statistical package.
Results
Herbaceous species composition
Comparison of observed(Sobs)and estimated(SChao2)species richness indicates that sampling completeness ranged between 64 and 91%(Table 1).Based on Sobs,a total of 61 species of herbaceous plants were recorded across the whole study site.Of these,37(~61%)were found both under and outside the canopies,whereas 11(18%)and 13(21%)were unique to under and or outside the canopies,respectively.When both under and outside the canopies were taken together,43(71%),39(64%)and 35(57%)species were recorded from the light,moderate and heavy grazing sites,respectively (Supplemental Table 1).These results show that the species richness of the herbaceous layer was 10%higher on the light grazing site than on the moderate grazing site,23%higher on the light grazing site than in the heavy grazing site,and 11%higher on the moderate grazing site than on the heavy grazing site.
Similarly,when data from both under and outside the canopies were pooled for each grazing site,26(47%)species were unique to one of the three sites,i.e.,6,17 and 3 species were unique to the light,moderate and heavy grazing sites, respectively (Supplemental Materials:Table S1).In the case of grazing categories,the results of the Chao2 estimator analysis also showed similar trends with the observed richness values:(1)species richness was 24%higher on the light grazing site than on the moderategrazing site;(2)33%higher on the light grazing site than on the heavy grazing site;and,(3)7%higher on the moderate grazing site than on the heavy grazing site.However,Sobswas 4%greater in the outside canopy compared to the under canopy,whereas SChao2was 11%greater under the canopy than outside(Table 1).
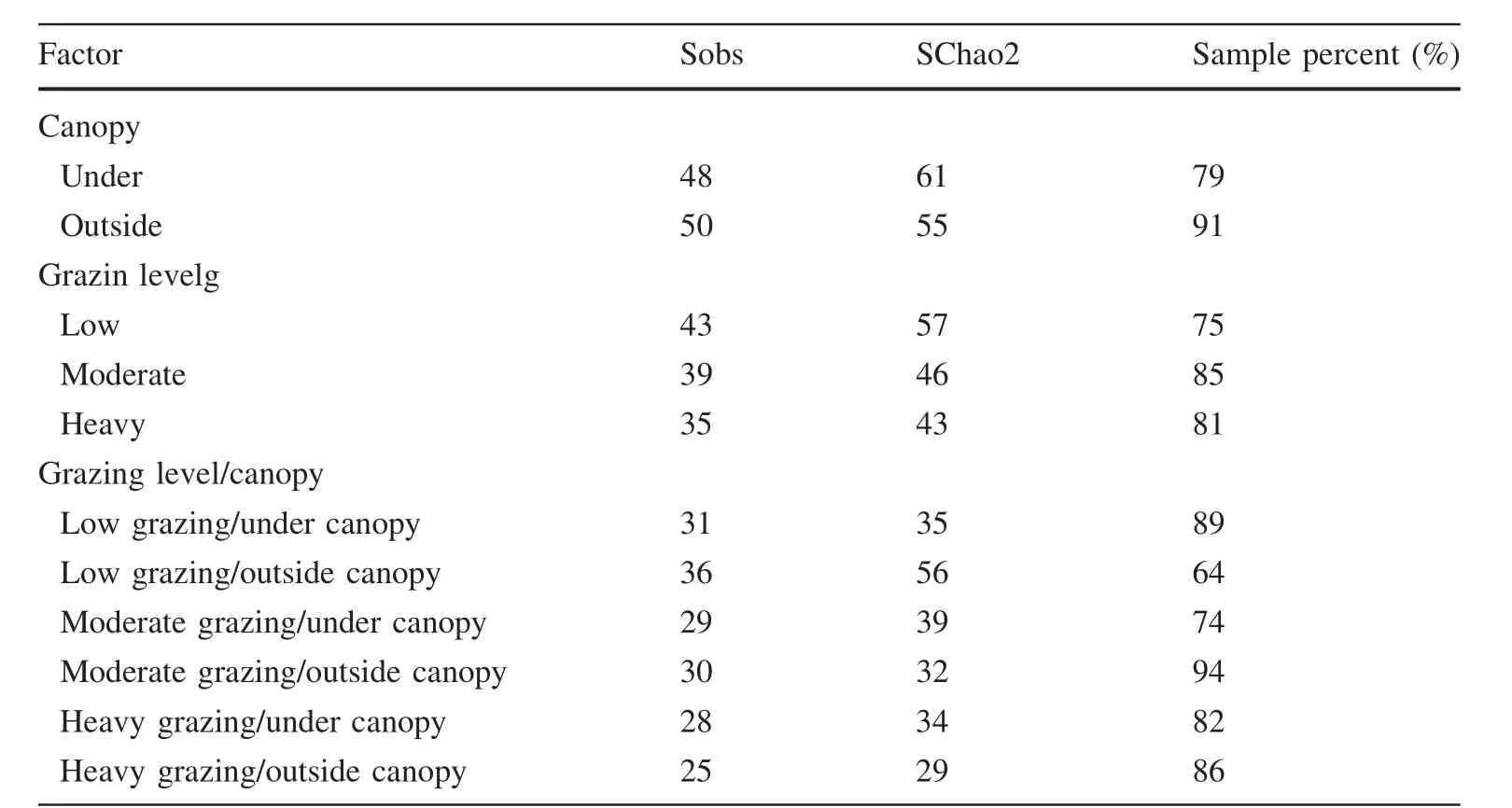
Table 1 Observed(Sobs)and Chao 2 estimated(SChao2)number of herbaceous species on sites of different factor levels
Regardless of the canopy effect,Sorensen’s index of similarity between the low and heavy grazing sites was 50%higher than that between the low and the moderate grazing sites and 31%between the heavy and moderate grazing sites(Table 2).The similarity index between under canopy and outside canopy vegetation was 75.5 when data from all sites were pooled,and ranged between 67.8(in the moderate grazing site)to 71.4(in the light grazing site)when each grazing level was treated separately(Table 2).The result of two-way analysis of similarity(ANOSIM)showed that both V.tortilis tree canopies(Global R=0.26,P<0.05)and grazing levels had significanteffects on herbaceous vegetation composition(Table 2,Fig.2).Considering the effects of V.tortilis’canopies in each grazing site separately,significant differences in species composition between under canopy and outside canopy were found only for moderate and heavy grazing sites(Table 2,Fig.2).

Table 2 Results of species compositional similarity analyses showing Sorensen’s similarity indexes(SI)and Global R values(R)between each possible pair of grazing and canopy treatment sites

Fig.2 Non-metric multidimensional scaling(MDS)ordination of samples from under and outside the canopy across three grazing(GR)gradients:grazing(GR)factors:LG lightly grazed,HG heavily grazed,MG moderately grazed;canopy factors:UC below canopy,OC outside canopy
Results of the SIMPER analysis showed that only four species,Achyranthes aspera L.,Justicia ladanoides Lam.,Sporobolus pyramidalis Beauv and Cenchrus ciliaris L.,contributed~20%of the dissimilarity between under canopy and outside canopy vegetation(Table S2a).Similarly,species such as Eragrostis aethiopica Chiov.and Chloris pycnothrix Trin.were more characteristic(i.e.,with higher percentage ground cover)of moderately grazed sites compared to lightly grazed sites,which in turn were characterized by Abutilon mauritianum(Jacq.)Medik(Table S2b).The vegetation of the lightly grazed sites was more differentiated against the heavily grazed sites by species such as Hyparrhenia rufa(Nees)Stapf.,Cynodon dactylon (L.) Pers. and Sida ovata Forssk.(Table S2c).Whereas,comparing the species composition of moderately and heavily grazed sites,the contribution to the dissimilarity of species like Hypoestes forskaolii(Vahl.)R.Br.,Eragrostis aethiopica and H.rufa appear to be higher(Table S2d).
Herbaceous biomass and basal cover
A significant effect of the canopies was revealed only on overall herbaceous biomass (grass plus herbs)(F1,66=6.294,P<0.05)and herb cover(F1,66=37.230,P<0.05)(Figs.3a,4b).Both were greater in the under canopy than in the outside canopy(Figs.3a,4b).Overall vegetation(F2,66=49.455,P<0.05)and grass biomass(F2,66=55.530,P<0.05)showed significant monotonic decline with increasing grazing pressure(i.e.light grazing>moderate grazing>heavy grazing) (Fig.3b).Although percentage ground cover of the overall herbaceous vegetation,grass and herbs in the low and heavy grazing sites were not significantly different,ground cover of the three vegetation groups were significantly greater in the moderate grazing sites compared to the low and heavy grazing sites (in all cases, F2,66=14.303–54.462,P<0.05;Fig.4b).
Soil variables and relationships with vegetation variables
Except for phosphorus,which showed no significant differences between treatments(F2,66=0.218,P=0.805),both canopies (F1,66=5.558–47.778,P<0.05) and grazing(F2,66=18.735–50.525,P<0.05)had significant effects on all soil variables.The three variables(moisture,carbon and nitrogen)were significantly greater under the canopies compared to outside the canopies(Fig.5a).Mean soil moisture content was similar both in the light and moderate grazing sites,but significantly lower in the heavy grazing site.However,the amount of carbon and nitrogen were similar on the light and heavy grazing sites,and significantly lower on the moderate grazing sites(Fig.5b).The results of multiple linear regression show that soil variables had significant relationships with all vegetation parameters(ANOVA:F4,67=3.110–17.047,P<0.05).However,model coefficients indicate that soil moisture had significant positive effects on overall vegetation and herb biomass,and negative effects on grass cover.Phosphorus had significant positive effects on herb biomass,while carbon had negative effects on herb cover(Table 3).
Discussion
Our results demonstrate that both V.tortilis canopies and grazing pressure have significant effects on herbaceous plant composition,biomass and cover.Although canopy had little effect on species richness,increased grazing intensity resulted in a considerable decline in species richness.Further,overall herbaceous biomass and herb cover were significantly greater below the tree canopy than outside,and showed monotonic decreasing trends as grazing pressures increased.These results are in agreement with studies of semi-arid savannas(Belsky et al.1993b;Jake and Michael 1990).
Trees influence plant species composition via several mechanisms.For example,they regulate the amounts of nutrients,moisture and light available to species growing below the canopy(Scholes and Archer 1997;Ludwig et al.2001;Asefa et al.2015a).Thus the probability of species occurrence under tree canopies depends on species-specific ecological/life history traits(e.g.,light-demanding vs.shade-tolerance)that determine their responses to these limiting factors(Scholes and Archer 1997;Ludwig et al.2001;2004b).However,such species responses could be modified by other ecological factors such as livestock grazing.In addition to their direct impact via selective grazing,vegetation removal and trampling,herbivory activities can affect herbaceous composition indirectly bychanging soil properties.As a result,grazing-tolerant plants are usually the most dominant groups on sites heavily disturbed by grazing,but disturbance-sensitive species tend to show negative responses to grazing intensities(Belsky et al.1993b;Dyer 2002;Asefa et al.2015a,b;Bikila et al.2016).Our findings of distinct vegetation composition between under canopy vs.outside canopy and among the three grazing levels support these ecological hypotheses.As shown by the SIMPER analysis,grass species such as E.aethiopica,Chloris pycnothrix,Cynodon dactylon and Sporobolus pyramidalis that are known to be indicators of land degradation(Abule et al.2005;Tessema et al.2011),were more abundant on the heavily grazed site compared to the light or moderate grazing sites.In contrast,species known to be indicators of healthy and little disturbed ecological sites of semi-arid savannas such as Hyparrhenia rufa,Heteropogon contortus and Cenchrus ciliaris(Tessema et al.2011,2012),appeared to be more abundant on lightly or moderately grazed sites.
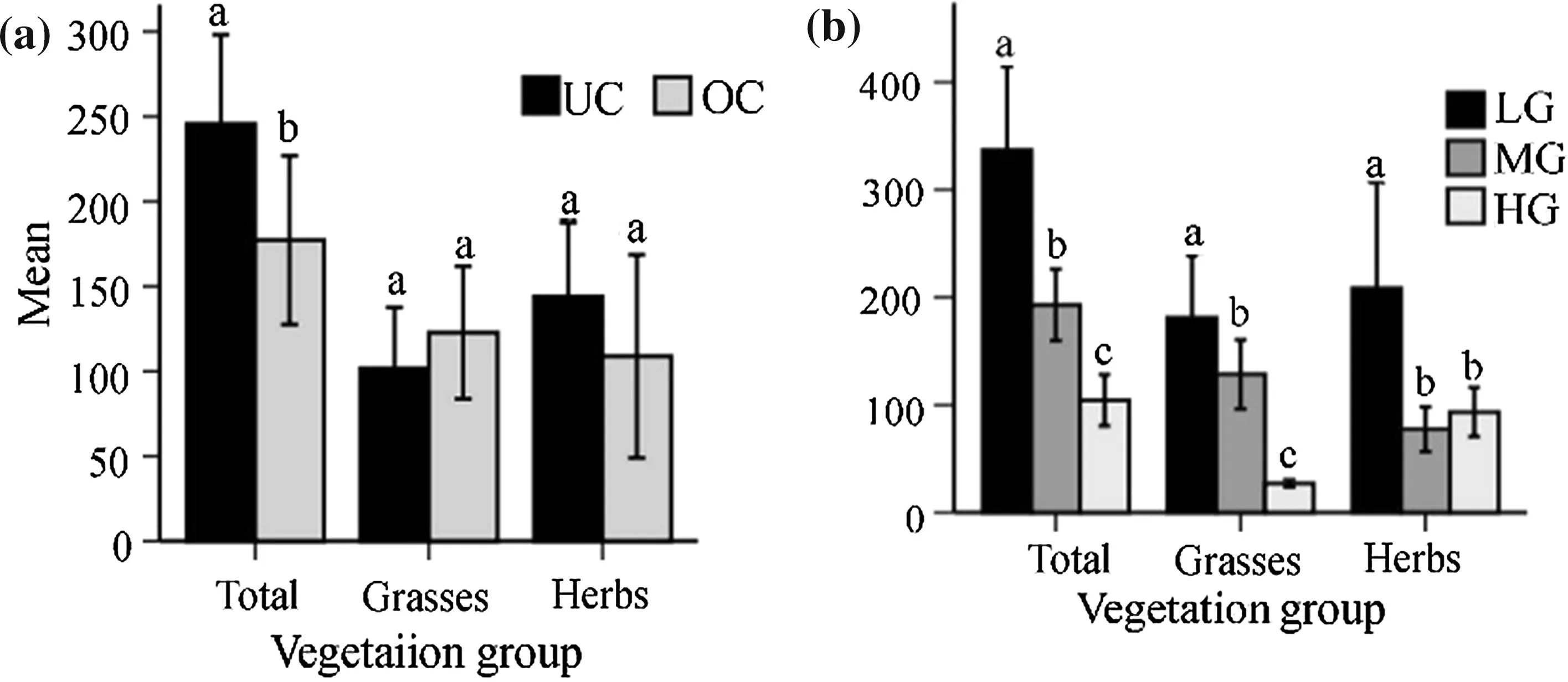
Fig.3 Mean(±SE)biomass of total(overall vegetation)grasses and herbs,a under the canopy(UC)and outside the canopy(OC),and b along grazing gradients(LG light grazing,MG moderate gazing,HG heavy grazing).Mean differences for each variable are indicated by different letters

Fig.4 Mean(±SE)ground cover of total vegetation,grasses and herbs,a in the under canopy(UC)and outside canopy(OC),and b along grazing gradients(LG light grazing,MG moderate gazing,HG heavy grazing).Mean differences for each variable are indicated by different letters
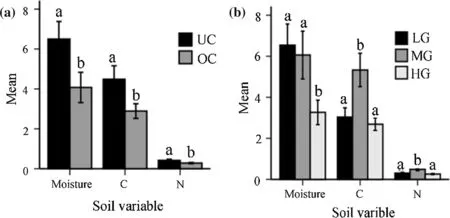
Fig.5 Mean(±SE)percentage of soil variables a in the under canopy(UC)and outside canopy(OC),and b along grazing gradients(LG light grazing,MG moderate gazing,HG heavy grazing).C carbon,N nitrogen.Phosphorous is not depicted as the values were too small.Mean differences for each variable are indicated by different letters

Table 3 Multiple linear model coefficients relating each vegetation variable with soil variables(coefficients only for soil variables that showed a significant relationship with vegetation variables)
Our results also indicate that in the central rift valley of Ethiopia,canopies of V.tortilis have positive effects on overall herbaceous biomass.This may be attributed to their effect on reducing soil moisture loss and increasing nutrient contents,as con firmed by our study.These variables appeared to have higher values below the canopy than outside the canopy and in particular,moisture content was positively correlated with vegetation biomass.The leaf area indices of our study trees are similar(Yadeta unpublished data),suggesting that the influence of differential light competition between canopy and understory vegetation is negligible.Similarly,Linstädter et al.(2016)reported that an intermediate canopy of tree legumes in a semi-arid African environment may increase species diversity and herbaceous biomass up to 32 and 29%,respectively,whereas,species with the most open canopies did not exert significant effects on the under story.Tree root facilitation rather than competition,and reduced evapotranspiration by canopy shading rather than light competition,might explain these findings.When herb and grass biomasses were examined separately however,the effect of canopy was insignificant on either.Although our results are affirmed by similar studies(Ludwig et al.2001,2004b)but other studies contradict our findings(Belsky et al.1993a;Abule et al.2005).
Two possibilities may explain inconsistencies about the effect of canopy on grass biomass:species composition and seasonality.First,highly productive,shade-tolerant grasses(i.e.,Panicum maximum Jacq.)are known to associate with trees resulting in increases in grass biomass in East African savanna biomes(Abule et al.2005;Angassa and Oba 2010).Such productive species were encountered less in our study,perhaps leading to a lack of differences between below versus outside canopies.Second,like most studies that support our findings(Belsky et al.1993b;Abule et al.2005;Dabasso et al.2014),our study was undertaken at the end of the rainy season,a period when most of the grasses were in flowering and seed setting stages.Thus,higher water and nutrient availability below the canopy might have been used for flowering and seed development than for vegetative components which in fluence the amount of biomass.However,this result should be interpreted cautiously as water may not be a limiting factor for the productivity difference observed.
Previous studies on the direct and indirect effects(e.g.,modifying soil physical and chemical properties)of grazing on vegetation in various ecosystems show in consistent results and suggest that the effects depend on,inter alia,the intensity of the grazing regime(Dyer 2002;Bradd et al.2011;Hailu et al.2015).In general,continuous heavy grazing leads to decreased diversity and biomass directly through vegetation removal,and hindrance of recruitment indirectly through soil compaction and modifying nutrient recycling processes(Hailu et al.2015;Asefa et al.2015a,b).However,grazing practices under moderate levels have been suggested as a management tool in semiarid savannas to increase vegetation diversity and biomass because they may lead to increased soil fertility(Abule et al.2005;Bradd et al.2011;Tessema et al.2011;Bikila et al.2016).Although,in contrast to our hypothesis,higher biomass was not demonstrated in the moderately grazed site of our study area,our findings of greater vegetation cover and increased soil nutrients(carbon and nitrogen)could be explained by this intermediate grazing hypothesis.Our results should be interpreted cautiously because there are a number of factors that may influence them,including arbitrarily de fining grazing intensity levels and size difference of study sites that may have confounding effects.Nevertheless,the effects of grazing activities,such as the selective removal of productive species that favor the growth of less productive,unpalatable species,might have contributed more to reduced biomass but increased cover in the moderate grazing site compared to the low grazing sites in our study.
Conclusion
The findings indicate that V.tortilis promotes herbaceous vegetation diversity and biomass by reducing soil moisture loss and increasing soil fertility.In contrast,increasing intensity of grazing has disastrous consequences to herbaceous vegetation diversity and biomass.This may best be explained to its influence on reducing soil moisture rather than to its influence on soil fertility.Furthermore,heavy grazing results in the shifting of highly productive perennials to less productive annual grasses,leading to changes in herbaceous composition and biomass.Therefore,we suggest that management practices should reduce the pressure on V.tortilis and that grazing should be regulated;low to moderate grazing levels(i.e.,a stocking rate less than by the Abernosa Cattle Breeding Ranch)appears to be tolerable to ensure sustainable conservation and use of the tree species in the study area in particular,and in semi-arid savannas in general.
AcknowledgementsWe would like to duly acknowledge the staffs of the Abernosa Cattle Breeding Ranch and of the Abijata Shalla Lakes National Park for their cooperation and assistance during the field data collection.We are also grateful to Ejigayeh Garanfa and Ferede Yadeta for their assistance in sample processing.Finally,the first author(TY)would like to express his heartedly thanks to the Wageningen University for his scholarship opportunity(Funding no:WUSRS-1205-10),and the Ethiopian Wildlife Conservation Authority for the study leave.
 Journal of Forestry Research2018年6期
Journal of Forestry Research2018年6期
- Journal of Forestry Research的其它文章
- Black locust(Robinia pseudoacacia L.)as a multi-purpose tree species in Hungary and Romania:a review
- The impact of the environmental factors on the photosynthetic activity of common pine(Pinus sylvestris)in spring and in autumn in the region of Eastern Siberia
- Osmoregulators in Hymenaea courbaril and Hymenaea stigonocarpa under water stress and rehydration
- Effect of nitrogen levels on photosynthetic parameters,morphological and chemical characters of saplings and trees in a temperate forest
- Free amino acid content in trunk,branches and branchlets of Araucaria angustifolia(Araucariaceae)
- Exogenous application of succinic acid enhances tolerance of Larix olgensis seedling to lead stress