
Effect of genetic sources on anatomical,morphological,and mechanical properties of 14-year-old genetically improved loblolly pine families from two sites in the southern United States
2018-11-06 06:44CharlesEssienBrianViaGiftyAcquahThomasGallagherTimothyMcDonaldLoriEckhardt
Journal of Forestry Research 2018年6期
Charles Essien•Brian K.Via•Gifty Acquah•Thomas Gallagher•Timothy McDonald•Lori Eckhardt
Abstract Tree improvement programs on loblolly pine(Pinus taeda)in the southeastern USA has focused primarily on improving growth,form,and disease tolerance.However,due to the recent reduction of design values for visually graded southern yellow pine lumber(including loblolly pine),attention has been drawn to the material quality of genetically improved loblolly pine.In this study,we used the time-of- flight(TOF)acoustic tool to assess the effect of genetic families on diameter,slenderness, fiber length,micro fibril angle(MFA),velocity and dynamic stiffness estimated using green density(DMOEG)and basic density(DMOEB)of 14-year-old loblolly pine stands selected from two sites.All the 184 and 204 trees of the selected eight half-sib genetic families on sites 1 and 2 respectively were tested using TOF acoustic tool,and two 5 mm core samples taken at breast height level(1.3 m)used to for the anatomical and physical properties analysis.The results indicated a significant positive linear relationship between dynamic MOEs(DMOEGand DMOEB)versus tree diameter,slenderness,and fiber length while dynamic MOEs negatively but nonsignificant correlated with MFA.While there was no significant difference in DMOEBbetween sites;velocity2for site 1 was significantly higher than site 2 but DMOEGwas higher for site 2 than site 1.Again,the mean DMOEGand DMOEBreported in the present study presents a snapshot of the expected static MOE for green and 12% moisture conditions respectively for loblolly pine.Furthermore,there were significant differences between families for most of the traits measured and this suggests that forest managers have the opportunity to select families that exhibit the desired fiber morphology for final product performance.Lastly,since the dynamic MOE based on green density(DMOEG),basic density(DMOEB)and velocity2present difference conclusions,practitioners of this type of acoustic technique should take care when extrapolating results across the sites.
Keywords Genetic families·Loblolly pine·Time-offlight·Resonance-based ·Modulus of elasticity ·Modulus of rupture
Introduction
In the southeastern U.S.,loblolly pine(Pinus taeda L.)is the most dominant tree species in the southern yellow pine(SYP)group in terms of cultivation,utilization and genetic improvement(McKeand et al.2003).One of the most important premium products from this species is dimensional lumber used for structural applications.There are several mechanical properties used to categorize solid wood and wood products.The two most widely measured for stiffness and strength are the modulus of elasticity(MOE)and modulus of rupture(MOR),respectively(FPL 2010).Isik et al.(2011)emphasized that MOE is one of the most important mechanical properties of wood and engineered wood products derived from loblolly pine.Consequently,in 2013 the design values for visually graded southern yellow pine lumber were adjusted in an attempt to reflect the material strength and stiffness of today’s market(ALSC 2013).For many product dimensions,these values dropped,making U.S.southern yellow pine lumber less competitive on the international market.In the southeastern USA,tree improvement programs on loblolly pine over the years focused primarily on improving growth,form,and disease tolerance with no consideration to material qualities such as stiffness and strength.However,due to this reduction in design values for visually graded southern yellow pine(including loblolly pine),attention has been drawn to material quality of genetically improved loblolly pine in our plantations.There is,therefore,a need to study the wood quality properties of plantation-grown trees.Traditionally,small,clear wood samples have been used to directly determine the effect of site,genetic source and planting density on MOE and MOR.For instance,Clark et al.(2008)studied the effect of planting density on MOR and MOE for loblolly pine using small clear samples and reported that these properties decreased with decreased planting density.MOE has been found to be highly or moderately heritable(Johnson and Gartner 2006;Lenz et al.2013),indicating that trees with a superior modulus of elasticity can be bred and deployed for plantation development.
Given that MOE is used to nondestructively classify lumber quality in the manufacturing plant using machine stress grading technology,estimation of MOE in both standing trees and logs would be of considerable use to tree breeders,mill managers,and silviculturists.For this purpose,several non-destructive instruments have been developed that can directly and/or indirectly measure the intrinsic wood quality of small samples,logs or trees(Wang et al.2001,2004;Mora et al.2009;Essien et al.2017b).Among these instruments are acoustic–based nondestructive tools which are simple,compact,and easy to operate.(Dickson et al.2004).Several researchers have found a strong linear relationship between the static MOE of small clear samples against the predicted dynamic MOE of small clear samples,logs and trees for different species(Wang et al.2001,2004;Lachenbruch et al.2010;Vikram et al.2011;Raymond et al.2008;Essien et al.2016,2017b).Currently,the most popular acoustic techniques are ‘‘time-of- flight’’and ‘‘resonance’’for standing trees and logs respectively(Wang et al.2001;Raymond et al.2008).
The time-of- flight(TOF)method measures the time it takes for an introduced stress wave to travel from point to point and currently is the only practical acoustic method for estimating the dynamic MOE in standing trees(Lasserre et al.2007;Wang 2013;Essien et al.2016).Therefore,TOF tools can be used for pre-harvest and progeny screening programs.Additionally,they can be used to predict the effect of edaphic,environmental,silvicultural and genetic factors on MOE of standing trees.This presents a unique opportunity for inclusion of stiffness screening in tree breeding programs.
Lasserre et al.(2007)studied the effect of genetic source,planting density and site on MOE of 11-year-old radiata pine.They observed MOE increased with greater planting density but there was a significant interaction between site and genetic source.Roth et al.(2007)observed similar results for the effect of planting density,genotype,and management operations on MOE for 6-yearold loblolly pine.Furthermore,Essien et al.(2016)reported thinning operations for loblolly pine at 13 and 22 years after planting significantly increased the MOE of standing trees by the 29th year.Essien et al.(2017a)attributed the acoustic signal to be sensitive to the same suite of fiber morphology and chemistry as a real stick of wood which is important if acoustics are to be used for the determination of stiffness in juvenile or younger wood.This is because Via et al.(2009)found MFA and lignin to be more relevant to stiffness in younger tissue,while density and cellulose were more important for older growth material.
Different authors have reported different trends for the relationship between velocity and dynamic stiffness against diameter at breast height(DBH)of trees.Lasserre et al.(2005)and Carson et al.(2014)noted a negative relationship between stiffness and DBH for 11-and 17-year-old radiata pine respectively.Briggs et al.(2007),on the other hand,found a positive relationship for Douglas- fir between 32-and 36-years-old while the relationship was negative for 42-year-old trees.The relationship between velocity and DBH was positive for the thinned stand while it was negative for the unthinned stand(Essien et al.2016).The relationship between velocity and stiffness against DBH appears to be affected by age,species and silvicultural operations(Wang et al.2004;Briggs et al.2007;Essien et al.2016).This divergent relationship between diameter and dynamic MOE estimated by TOF might be due to the fact that the generated sound waves are restricted to the sapwood portion that is penetrated by the probes.Since fast-growing species produce wood with thin and large lumen cells,one would expect a lower velocity and stiffness.Furthermore,the velocity of fast-growing trees will be reduced due to a higher percentage of free water in the cell lumen.
Since loblolly pine constitutes the major part of SYP,understanding the main and interactive effects of available planting stock and site in relation to MOE is of utmost importance for the growth and sustainability of the forest industry.If statistically,significant relationships exist between the main and interactive effects of site and genetic sources,then a predictive model for loblolly pine must consider the site and genetic sources of the materials.We hypothesize that there will be a significant site and genetic sources effect on the diameter,velocity,and dynamic MOE.Therefore,the main objective of this study is to determine the main and interactive effects of genetic sources on physical,anatomical and quality properties of fifteen genetically improved,half-sib loblolly pine families selected from two sites.
Materials and methods
Trees were selected from a plantation of genetically improved,open-pollinated half-sib families of loblolly pine established at two sites in the southeastern U.S.in 2000.Site 1 was in Nassau County,Florida near Yulee(latitude 30°63′N and longitude 81°57′W),and Site 2 at Brantley County,Georgia near Nahunta(latitude 31°12′16′N and longitude 81°58′56′W.Soils of Site 1 are poorly drained and formed from a thick bed of alkaline loamy and clayey marine sediment with slopes less than 1%.The mean longterm temperature and precipitation from 1981 to 2010 are 21°C and 1350 mm respectively.The topography of Site 2 is relatively flat with a slope less than 2%and 20 masl.Soils are fine sandy loams,poorly drained and generally poor in nutrients,and formed from loamy and silty Coastal Plain sediments.Mean annual temperatures range 17–19 °C and annual precipitation from 1981 to 2010 averaged 1315 mm(NOAA 2016).
A randomized complete block design was used on both sites.Site 1 was 6744 m2and site 2 8027 m2and each site was divided into fifteen blocks measuring 450 and 535 m2respectively.A single tree per block design was used with 80 seedlings from eighty different genetically improved half-sib families randomly planted at 1.6 m×3.2 m and 1.8 m×3.6 m for site 1 and 2,respectively.The stands had not been commercially thinned since establishment at the time of data collection.
Fifteen out of the eighty loblolly pine families were selected a priori based upon their tolerance to fusiform rust(Cronartium quercum(Berk.)Miyabe ex Shirai f.sp.fusiforme)and tested on both sites in the spring of 2014 when the trees were 14-years-old.Although 15 seedlings per genetic family were planted,seedling mortality over time reduced the number of trees in some families.In all,388 trees comprising 184 and 204 for sites 1 and 2,respectively,were assessed.Heights and diameters were measured using clinometer and diameter tape.
Acoustic measurements of standing trees
All of the trees from the selected fifteen families were acoustically tested using the FAKOPP Microsecond Timer acoustic tool,(Fakopp Enterprise,Agfalva,Hungary),which relied on the time-of- flight(TOF)principle(Wang et al.2007;Mora et al.2009;Essien et al.2016).The accelerometers(transmitter and receiver probes)were positioned on the same side of the tree 120 cm apart with the center of the path at breast height.Both probes were positioned 45°to the tree axis and the stress wave was generated by striking the transmitter probe with a steel hammer at a steady force(Mora et al.2009).The generated wave was detected by the receiver and the time lapse for the sound wave to travel the distance between the probes was recorded by the data logger.Seven readings were taken on each tree and velocity was estimated as the ratio of the distance to time(Eq.1).The dynamic modulus of elasticity estimated with green density(DMOEG)used Eq.2(Wang et al.2007;Essien et al.2016).However,acoustic velocity has been found to decrease with increasing moisture content until the fiber saturation point,above which changes in moisture insignificantly affect velocity(Olivito 1996;Chan et al.2011;Gao et al.2012).Since the moisture in healthy trees,including loblolly pine,is above the fiber saturation point,we assumed that velocity will be constant hence basic density was used to estimate the dynamic MOE of the tree(DMOEB)using Eq.3.Basic density is the ratio of the oven-dry weight to the green volume of a piece of wood(FPL 2010).Given that the oven-dry weight is the smallest constant weight of the material and the green volume is the largest volume,basic density is the smallest possible density of wood.Therefore,using basic density and velocity should provide important information about wood quality.Two 5-mm thick core samples were taken at breast height of each tree to determine moisture content,basic density,green density,and anatomical properties.One of the 5-mm samples meant for moisture content and densities determination was cut into 2-cm long segments,yielding 3–12 wood segments.The dimensions of the segments were measured using a digital caliper to the nearest 0.0025 mm and weight to the nearest 0.001 g following the procedure described in ASTM D2395-2007.The diameter of the segmented cores was measured at both ends and the average used for volume determination.A digital caliper was used to determine the volume of the core samples because,according to Via(1998),the displaced determined volume was highly correlated with the caliper determined volume(R2=0.99).However,3–12 replicates were used to reduce potential errors in volume determination.The average of all the segments from the same core was used for analysis.Weight(Mi)and dimensions of the samples were measured while green and were subsequently dried to constant weight in an electronic oven at 103°C(Mo).The moisture content(MC),basic(ρbasic)and green densities(ρgreen)of the cores were determined according to Eqs.4,5,and 6 respectively.

where VTis tree velocity in m/s,D the distance between the probes in meters,s the time for the stress wave to travel the distance between the probes in seconds,DMOEGthe dynamic modulus of elasticity estimated by green density,DMOEBthe dynamic modulus of elasticity estimated by basic density.MC is green moisture content of the core,Mithe initial green weight,Mois the oven dried weight,ρgreenis the green density of the core,Vgreenis the green volume of the core.
MFA and fiber geometry determination
Subsamples of eight families were selected from the fifteen families used for the study.The 15 families were ranked based on DMOEG for each and the combined sites.However,most the families ranked differently on each site,hence the subsampling was based on the combined site.Two families from the mid-rank(T34 and T37)while three families each from the higher(T18,T26,and T33)and lower(T3,T15,and T21)stiffness groups were selected for the anatomical study.Thin sections of samples measuring 20 μm were sliced from along the entire length of the samples from each family using a sliding microtome(Yamato Kohki ROM 380,Japan).The samples were macerated using equal volumes of hydrogen peroxide(30.0%)and glacial acetic acid(99.6%)at 80°C for 24 h(Peter et al.2003;Essien 2011;Essien et al.2017a).The macerated cells were rinsed thoroughly with deionized water and the fibers left in deionized water for 48 h,after which it was shaken rigorously prior to measurement.This was to ensure that the fibers separated for easy visualization.Slides were prepared from the macerated fibers and fiber length,diameter,and thickness measured on forty full fibers per family with 20 each selected from the earlywood and latewood using Differential Interference Contrast(DIC)Microscope(Olympus BX53,Janpan)(Essien 2011;Essien et al.2017a)and the digital images of the MFA taken using the DIC microscope(Peter et al.2003).The ImageJ software was used to measure the MFA.
Data analysis
Standing tree variables were DBH,height,slenderness(ratio of total height to DBH),velocity(VT)and dynamic modulus of elasticity(DMOEGand DMOEB).Pearson’s correlation coefficients were used to determine the level of linearity among the variables.The standing tree variables were analyzed using a generalized linear model(GLM)procedure in SAS 9.4(SAS Institute 2014)to determine the effect of families,block,site and their interactions on diameter,velocity and dynamic MOEs(DMOEGand DMOEB).If site×family interaction was significant at 5%,it indicated that slopes of the model were different and thus the two populations were different and were analyzed separately.However,if the interaction was not significant,then the two populations were similar and therefore they could be combined for analysis.In order to account for diameter and ratio of height to diameter(slenderness),variability effect on velocity,DMOEGand DMOEB,they were introduced as a covariate in the model(8).The covariates were assessed the effect of site and family on velocity and dynamic modulus of elasticity.

where Y is DBH,slenderness,tree velocity,micro fibril angle, fiber length or tree dynamic modulus of elasticity(DMOEGand DMOEB),β1is the mean change in Y due to site(site 1=1 and site 2=0),β2is the mean change in Y due to genetic family,β3,and β4is the mean change in Ydue to block and site×family interaction term,e is the residue.
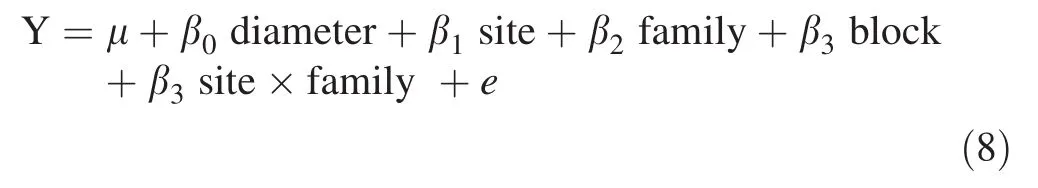
where Y is tree velocity(VT)or tree dynamic modulus of elasticity(DMOEGand DMOEB),β0is mean change in Y due to DBH or slenderness(taper).If the interaction term was significant at 0.05,then the slopes of the two sites are statistically different from zero.In such cases,least square means are generated for the fixed main parameter and the multiple comparisons performed using Tukey–Kramer adjustment at 0.05.
Results and discussion
Relationships between anatomical,morphological,and mechanical properties
There was a significant positive linear relationship between diameter and the two quality parameters,but diameter was negatively related to slenderness(Table 1).The negative linear relationship between diameter and slenderness(R=-0.129)was expected since slenderness is the ratio of the total tree height to diameter(Roth et al.2007).This con firmed previous studies where a significant positive relationship existed between standing tree stiffness and stem slenderness(Roth et al.2007;Lasserre et al.2007;Antony et al.2012).
Wood is an anisotropic material with most properties varying in radial,tangential,and longitudinal directions.Modulus of elasticity as one of the major mechanical properties of wood varied radially and axially as a result of the interactive effect of density and micro fibril angle(Megraw et al.1999;Via et al.2009).Depending on the age,between 76 to 96%and 66 to 94%of the variation in MOE of loblolly and longleaf pines,respectively,could be explained by density and MFA.Generally density has a positive correlation with dynamic MOE (R=0.75,R=0.42 for green and basic densities respectively)while increasing MFA reduces DMOEB(-0.13)(Table 1).Via et al.(2009)explained that,in the mature wood zone,density becomes a very important predictor of MOE since the magnitude of MFA becomes relatively small and stabilizes in this zone,while a high MFA with considerable variation assumes more influence in the core wood zone.This iteration leads to the formation of stiffer wood material in the outer wood than in the core wood zones.However,this is affected by the edaphic,environmental and silvicultural practices of the stands.Consequently,if the plantation is established at a high planting density,taller and thinner trees are produced due to the high intertree competition for light at the juvenile stage.Therefore,the core wood attains stable and small MFA earlier as compared to those established at low planting density,leading to increased MOE with slenderness as reported in this study.

Table 1 Pearson correlation coefficients among morphological,anatomical and quality parameters
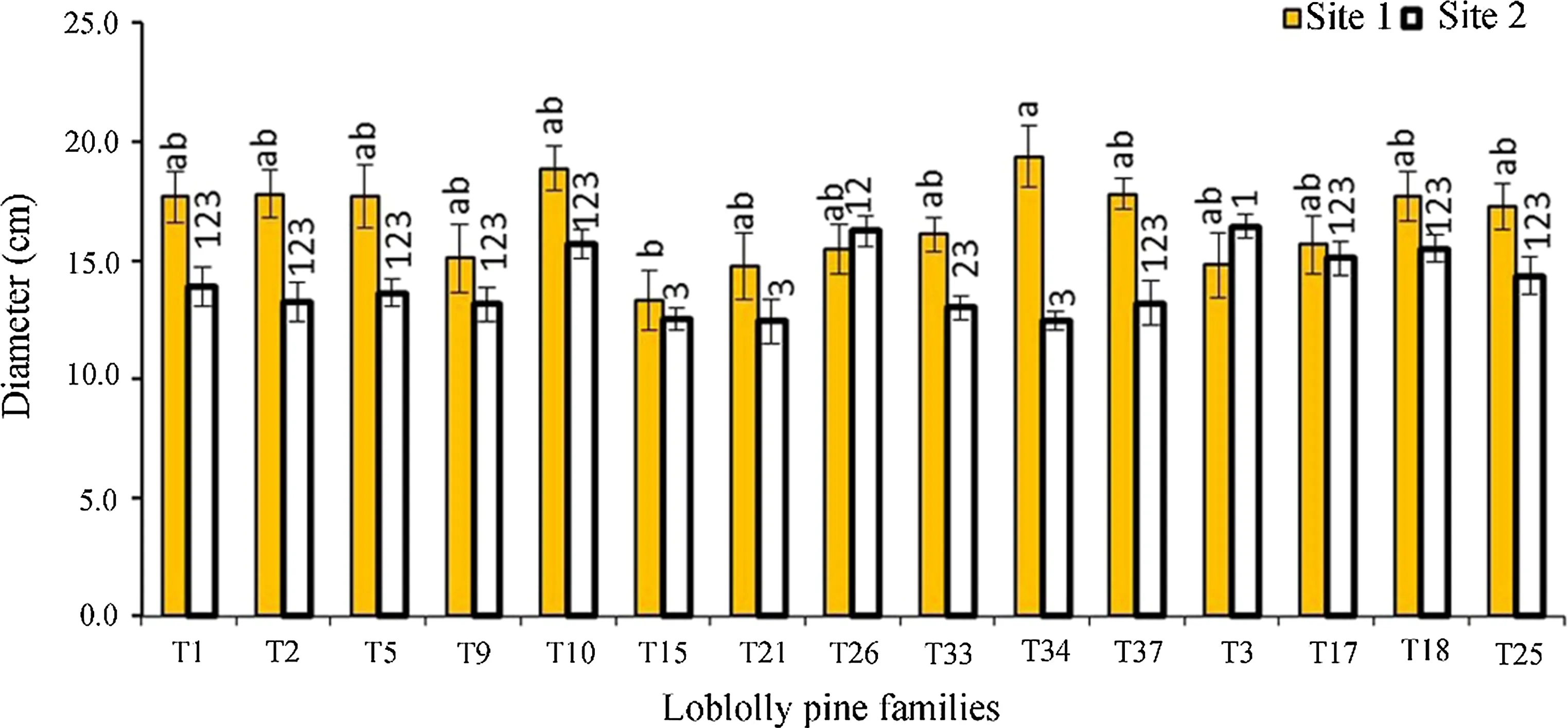
Fig.1 Comparison of mean diameter growth of families for each site.The error bars are standard deviations of an individual family.Families with the same letter or number are not significantly different at 0.05.**A significant difference between sites
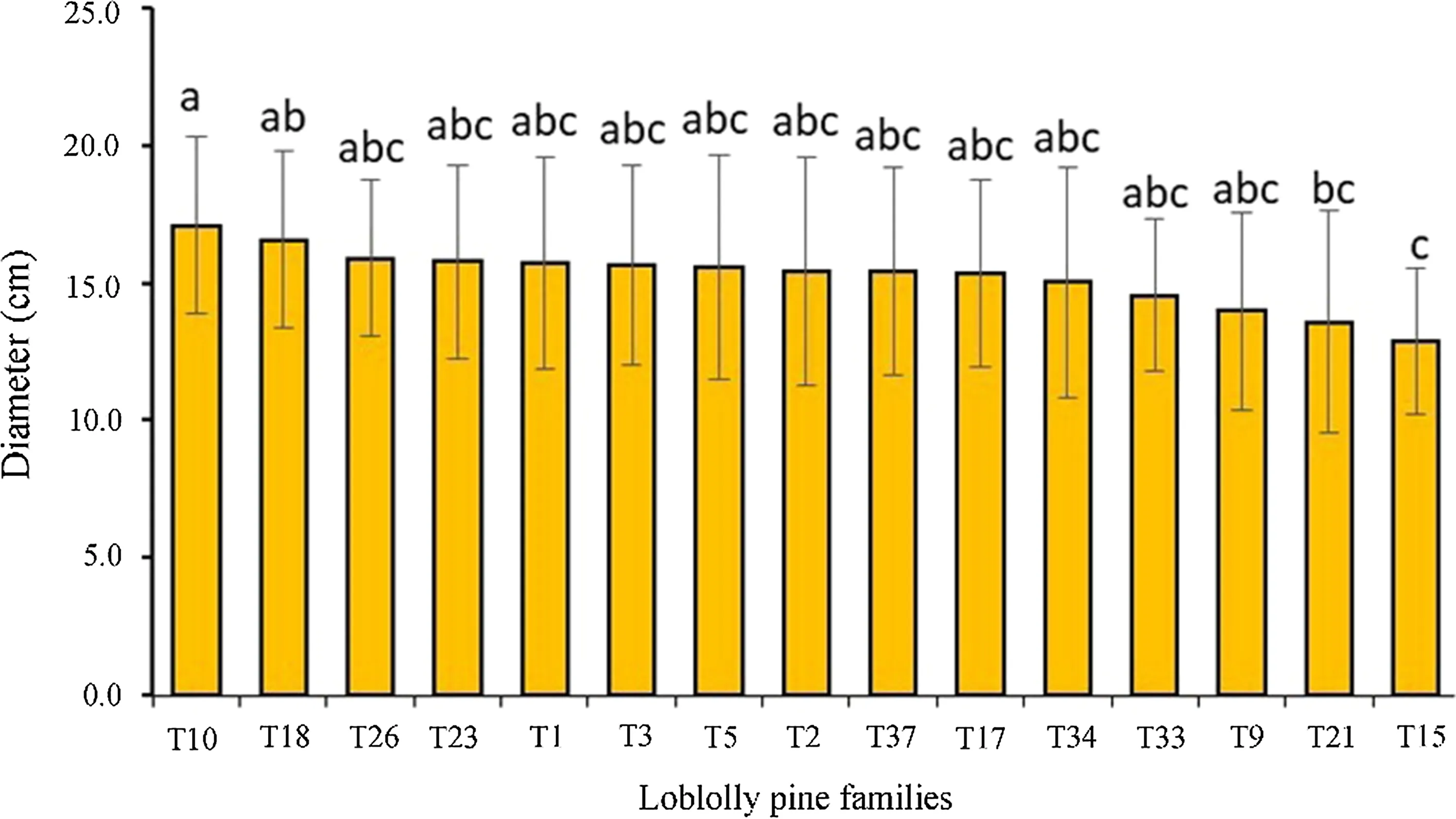
Fig.2 Ranks of mean diameter growth of families for the combined sites(n=388).The error bars are standard deviations of an individual family.Families with the same letter are not significantly different at 0.05

Fig.3 Comparison of mean velocity of families for each site.The error bars are standard errors of an individual family.Families with the same letter or number are not significantly different at 0.05

Fig.4 Ranks of mean velocity of families for the combined sites(n=388).The error bars are standard deviations of an individual family.Families with the same letter are not significantly different at 0.05
The positive relationships between the dynamic MOEs(DMOEGand DMOEB)and slenderness con firm the theory that a slender tree requires higher stiffness to support itself from buckling under its own weight(Greenhill 1918).Fundamentally,a taller tree is subject to higher wind load and the longer distance to the bottom causes a higher moment.Therefore,the tree requires stiffer material to withstand wind loading.


Table 3 Analysis of variance of the main effect of site,genetic sources and block on anatomical,morphological,and quality properties of 388 trees from both sites studied
However, correlations between MFA, velocity(R=-0.07),and DMOEG(R=-0.09)are nonsignificant as shown in Table 1.This result is unexpected,as it has been reported that a decrease in MFA will result in an increase in velocity because sound waves travel through the axis of the cellulose along the MFA(Hasegawa et al.2011).This may be due to the sampling method used in this study.Most studies investigating the effect of MFA on other wood properties optimize the range of MFA values through sampling,separating the mature outer wood from the juvenile core(34–63 years),such that MFA values cover a wider range(Via et al.2009;Hasegawa et al.2011).In this study,the random natural MFA range present in the 14-year-old samples were used,and there was no intention of selecting a wide MFA range to optimize its variance.
The positive relationship between the dynamic MOEs and diameter has been reported by several authors(Wang et al.2004;Briggs et al.2007;Essien et al.2016).However,other studies such as Lasserre et al.(2005)and Carson et al.(2014)reported a negative relationship between stiffness and diameter of 11-and 15-year-old Pinus radiata D.Don at different planting densities.Since MOE is moderately or highly heritable(Lenz et al.2013),diameter is not genetically correlated with MOE(Li et al.2007).Therefore,it is possible for a large diameter tree to produce stiff wood,and at the same time,for a large diameter to produce less stiff wood.An individual family by family analysis for diameter(Figs.1,2),velocity(Figs.3,4)and dynamic MOEs show families that,(e.g.,T26 and T18)have higher diameter,velocity and dynamic MOEs.Roth et al.(2007)made a similar observation when they studied 6-year-old elite loblolly pine.
Effect of site and genetic families on anatomical,morphological,and mechanical properties
The descriptive statistics of the parameters studied are in Table 2.All the parameters studied,with the exception DMOEBand fiber double-wall thickness,varied significantly with the site(p<0.05).Though the mean velocity averaged across the family was significantly higher for site 1,DMOEG,MFA,slenderness,and fiber diameter were larger on site 2(Table 2).
This is interesting because,based on Eqs.2 and 3,one would expect the site with higher velocity(Figs.3,4)to exhibit a higher dynamic MOE,since the higher order of velocity dominates the variation in the response variable for the equation.However,density,which is a function of fiber diameter and wall thickness on site 2,was higher than on site 1(Table 2).Consequently,both green and basic densities of trees on site 2 will be higher than on site 1 due to the large fiber lumen(for potential space for free water which inflates the weight of the wood)and the thicker wall thickness,respectively,hence causing the dynamic MOE to lean towards site 2.Therefore,using velocity2(Roth et al.2007),dynamic MOE based on basic density(DMOEB)and based on green density(DMOEG),lead to different conclusions.While there is no significant difference in DMOEBbetween sites,velocity2for site 1 is significantly higher than for site 2,but DMOEGis higher for site 2 than site 1.Therefore,practitioners should take care whenextrapolating velocity readings of trees from different locations.

Table 4 Mean(GPa)and standard error(SE)(GPa)of dynamic MOE(DMOEG)for each of the 15 genetic families per site for 388 trees
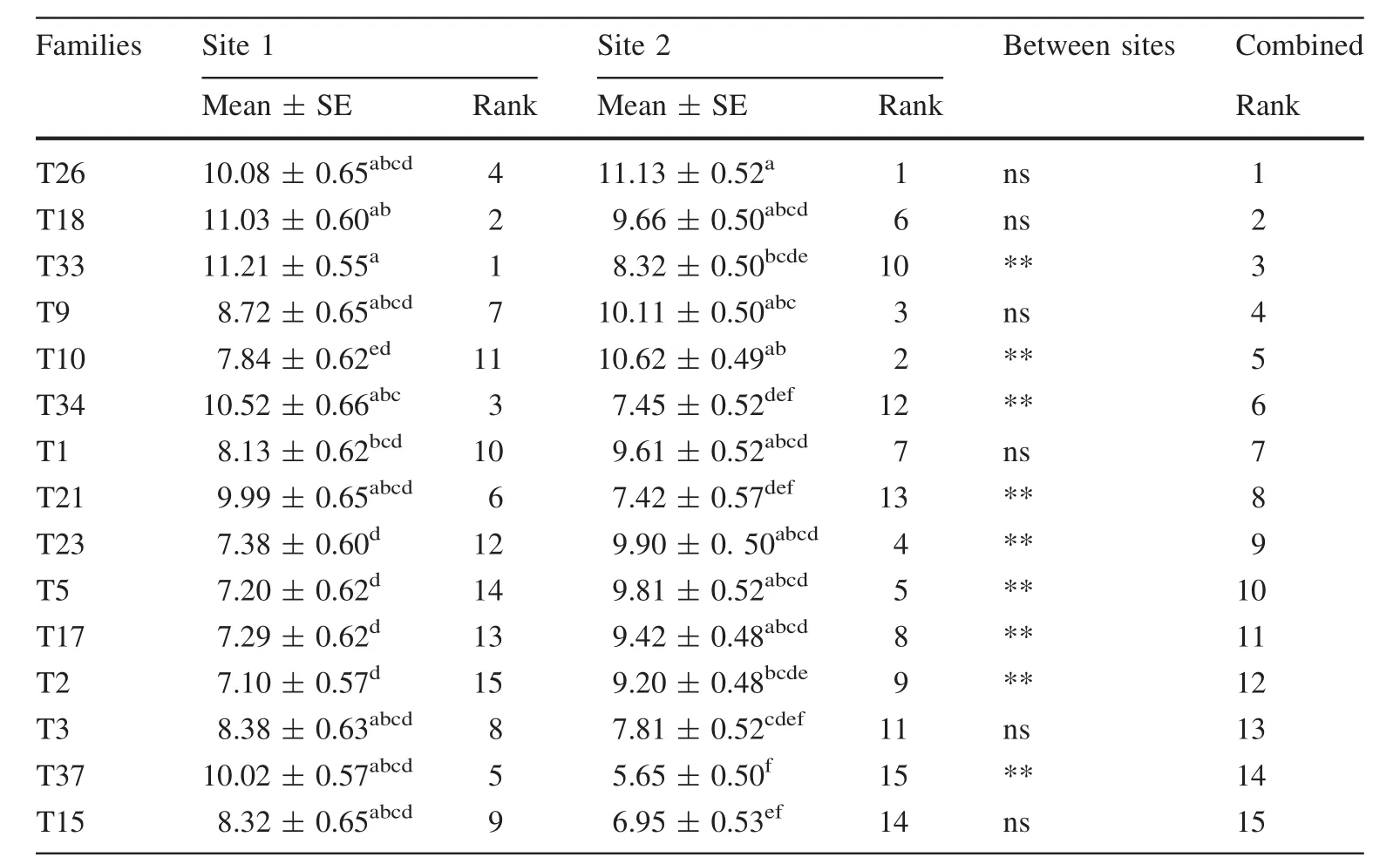
Table 5 Mean(GPa),standard error(SE)(GPa),and rank of dynamic MOE(DMOEB)for each of the 15 genetic families per site
FPL(2010)reported a mean static MOE for loblolly pine at green and 12%moisture conditions of 9.7 GPa and 12.7 GPa,respectively.This suggests that the mean DMOEBand DMOEGin the present study are very similar to static MOE for green and 12%moisture conditions,respectively.Generally,the analysis of variance indicates a significant(p<0.05)effect of site and families on tree size and form,MFA, fiber length,velocity,and dynamic MOEs(DMOEGand DMOEB) (Table 3, Fig.3). The site×family interaction was significant for the tree size,form,and dynamic MOEs(Table 3).This was mainly due to the changing in ranks of some families between sites.For example,half the families studied changed ranks in dynamic MOEs(Tables 4,5)and fiber diameter(Table 6).On the other hand,families 3 and 34 were the main causes of the significant interaction term observed for diameter(Fig.1),MFA(Table 7)and fiber length(Table 8).After adjusting for slenderness(ratio of height to diameter),neither the main effects nor the interaction significantly(p>0.05)affects velocity(Table 9).

Table 6 Mean(μm),standard error(SE)(μm),and rank of fiber diameter for each of the 8 subsampled genetic families and combined per site
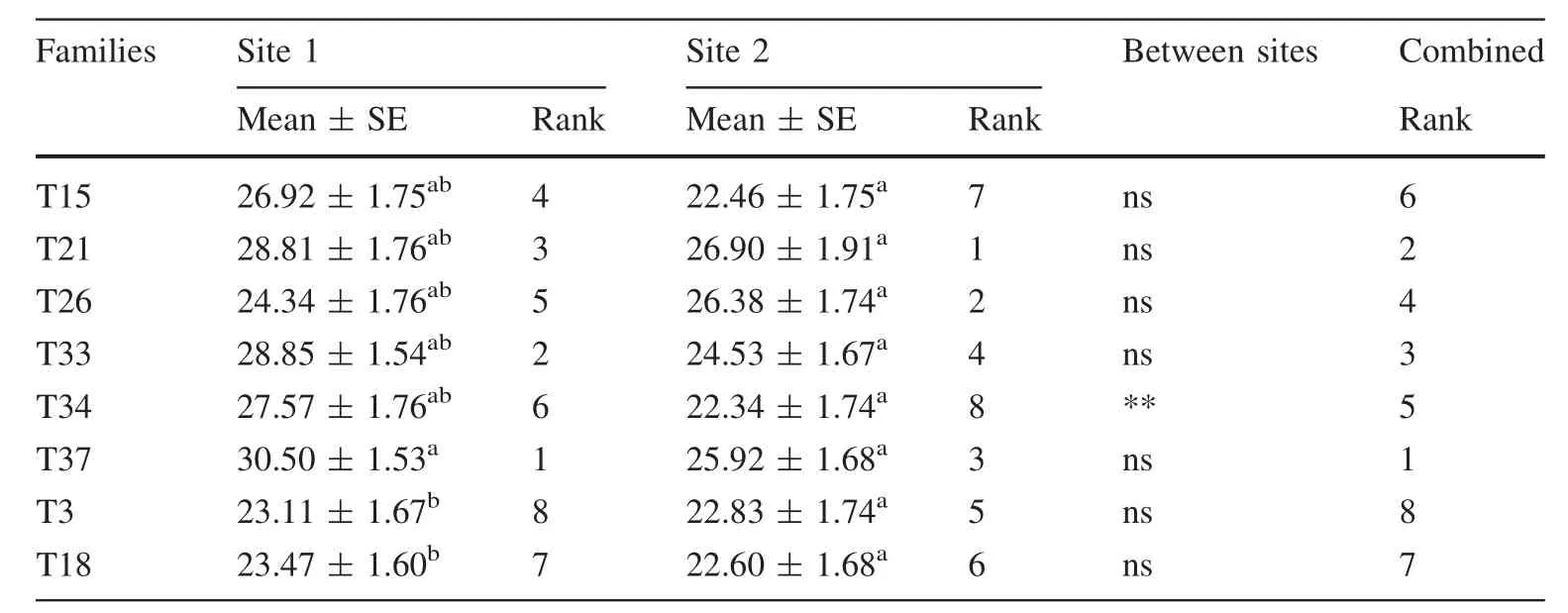
Table 7 Mean(°),standard error(SE)(°),and rank of fiber diameter for each of the 8 subsampled genetic families and combined per site

Table 9 Analysis of variance of the main effect of site,genetic sources and block on anatomical,morphological and quality properties of 388 trees after adjusting for diameter and slenderness
However,when similar adjustments were performed on the dynamic MOEs,both the main and the interaction between site×families were significant(Table 9).The stiffness of juvenile pines is significantly influenced by genetics(Dungey et al.2007)and edaphic conditions(Lasserre et al.2007).In this study,the significant main and interactive effects of site and families of dynamic MOEs persist,even after adjusting for slenderness.This indicates a strong effect of site and families on MOEs.
Site selection is one of the important management decisions in plantation establishment because,besides its effect on growth,form,and yields,site also influences the pest and disease management systems to use.For instance,for a given family on different sites,a fertile,well-aerated and drained site will produce larger,longer and healthy trees whereas those planted on less fertile and compact fields will be smaller,thinned crown and eventually show dieback.Generally,most of the families shared similar morphological and internal quality values,indicating the stability of the planting stock across the two sites.Therefore,there is a good potential for landowners and foresters to select families with superior wood qualities and fast growing performance for plantation establishment.However,for site sensitive families,resource managers should allocate the appropriate family to the require site for desired wood quality and growth performance.
Conclusions
The effect of site and genetic families on morphological,anatomical and wood quality properties of fifteen 14-yearold loblolly pines stock were studied The results indicate significant positive linear relations between dynamic MOEs(DMOEGand DMOEB)versus diameter,slenderness,and fiber length,while negatively and non-significantly correlated with MFA.
Furthermore,while there are no significant differences in DMOEBbetween sites,velocity2for site 1 is significantly higher than for site 2 but DMOEGis higher for site 2 than site 1.Therefore resource managers should take care when extrapolating velocity readings from different locations.The mean DMOEBand DMOEGreported in the present study presents a snapshot of the expected static MOE for green and 12%moisture conditions,respectively.
Additionally,50%of the selected families had statistically similar morphological and quality properties between the sites,suggesting that half the planting stock exhibit stability and homogeneity.Therefore,farmers and resource managers have the opportunity to select families which are superior in both the morphological and quality traits for plantation development across sites with varying soils and environments.Furthermore,there is the opportunity to place elite planting stocks to specific site for greater productivity and quality output due to the significant effect of site by family interaction.
AcknowledgementsThe materials for the study were supplied by the Auburn University Forest Health Cooperation.We appreciated the technical and logistical support from students and staff of Forest Health Dynamics Laboratory and Forest Products Development Center of Auburn University.
 Journal of Forestry Research2018年6期
Journal of Forestry Research2018年6期
- Journal of Forestry Research的其它文章
- Black locust(Robinia pseudoacacia L.)as a multi-purpose tree species in Hungary and Romania:a review
- The impact of the environmental factors on the photosynthetic activity of common pine(Pinus sylvestris)in spring and in autumn in the region of Eastern Siberia
- Osmoregulators in Hymenaea courbaril and Hymenaea stigonocarpa under water stress and rehydration
- Effect of nitrogen levels on photosynthetic parameters,morphological and chemical characters of saplings and trees in a temperate forest
- Free amino acid content in trunk,branches and branchlets of Araucaria angustifolia(Araucariaceae)
- Exogenous application of succinic acid enhances tolerance of Larix olgensis seedling to lead stress