
东海中南部日本鲭种群分析
2018-10-26 08:06刘尊雷马春艳严利平李圣法李建生
海洋渔业 2018年5期
刘尊雷,马春艳,严利平,刘 勇,李圣法,李建生,张 辉
(中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,上海 200090)
日本鲭(Scomber japonicus)属大洋暖水性中上层鱼类,广泛分布于西北太平洋沿岸海域,是我国东、黄海中上层最重要的渔业资源之一,主要由中国、日本、韩国等国家捕捞利用[1]。我国主要的捕捞方式为20世纪70年代发展起来的国有机轮灯光围网作业、群众机帆船灯光围网,还有少量的拖网兼捕作业[2]。随着捕捞规模不断增加和作业渔场的拓展,至21世纪初,我国东海区日本鲭的产量在20×104t左右波动,黄海区的日本鲭产量也从20世纪80年代的3×104t左右,提高到本世纪初的(11~12)×104t水平,已成为主要的经济鱼种之一,在我国的海洋渔业中占有重要的地位[3]。
种群是物种存在、生物遗传、进化、种间关系的基本单元,是生物群落和生态系统的基本结构单元,同时也是资源开发利用和管理的具体对象[4],种群鉴定的研究目的在于获取管理单元,借此减少评估模型中的不确定性和改善资源的管理现状[5]。只有在正确区分种群的基础上,了解鱼类种群结构才能为鱼类资源的合理利用和管理提供科学依据[4]。就东黄海的日本鲭种群鉴定和划分而言,中国和日本学者曾在20世纪50年代和70年代通过大量的标志放流并结合渔业调查数据研判了日本鲭的种群结构[6-8],认为分布于东海海域的日本鲭存在一个种群(东海群系)和两个种群(东海西部种群和五岛西部种群)的不同观点[9]。至21世纪,一些研究者基于前人研究结果,利用现代研究方法进行了相关验证[10-12],初步支持东、黄海的日本鲭存在东海西部和五岛西部的两个种群的观点[12]。随着2000年6月1日《中日渔业协定》的正式生效,中日渔业协定暂定措施水域重要渔业资源的配额分配等问题渐受关注,日本鲭作为该区域内的主要洄游分布种尤其受到高度重视。同时,日本大型灯光围网常年在东海东南部200 m等深线附近海域围捕中上层鱼类,而我国自1998年始大型围网逐年加强了对东海中南部海域的日本鲭利用,产量已超过黄海中南部[3],在该海域日本鲭的渔业优势日渐突出。鉴于上述原因,本研究主要针对包含中日暂定措施水域在内的东海中南部日本鲭进行区域性的种群判别,旨在为该海域日本鲭资源评价和管理提供科学依据。
1 材料与方法
研究样本取自2012-2016年2-5月东海中南部(26°15′~29°15′N、122°45′~126°45′E)大中型围网捕获的日本鲭,样本采集点共11个,其中,采集点③~⑩及※分布于中日渔业协定暂定措施水域的南部区域,采集点①和②分布于中日渔业协定暂定措施水域以南邻近区域(图1),计15批次,每批次的样本量在30~50尾,合计544尾,其体长范围为235~375 mm,性腺成熟度为Ⅲ~Ⅵ期。

图1 2012-2016年2-5月日本鲭样本采样点分布Fig.1 Sampling stations of Scomber japonicus during February-M ay 2012-2016
1.1 外部形态
外部形态分计数和度量两类性状,共分析10个采样点计14个批次。计数性状包括鳃耙数、胸鳍数、第一背鳍数、第二背鳍数和臀鳍数5个指标;度量性状选取10个定位点,对每尾样品测定包含一个叉长数据和20个框架数据[9],利用相对生长测定法(allometric method)对每个样品的原始框架数据进行标准化。相对生长测定法公式如下:

式中:Ladj为标准化后的框架值;L为原始框架数据;Fs为样本组的平均叉长;F0为每个样本的叉长,b为每个样本的log L对log F0的斜率。
利用典型对应分析法(canonical correspondence analysis,CCA)解析采样点之间群体计数性状的相关性,利用冗余分析法(redundancy analysis,RDA)解析采样点之间群体度量性状的相关性,上述方法中均以采样点为解释变量,并使用置换检验(permutation test)反复审定“所有采样点之间群体性状没有任何差异”这一虚假设。统计分析使用R语言(3.01版本)的 vegan包(2.2-1版本)。
1.2 分子遗传学
采用DNA分子标记共分析8个采样点9个批次的样品,对每采样点取20~30尾日本鲭尾部肌肉(100 mg·尾-1),按 SAMBROOK等[13]方法提取基因组DNA,mtDNA Cytb基因使用通用引物 L14724(5′-GAC TTG AAA AAC CAC CGT TG-3′)和 H15915(5′-CTC CGA TCT CCG GAT TAC AAG AC-3′)扩增而获得。PCR产物用UNIQ-10(上海生工)柱式纯化试剂盒纯化,纯化产物在ABI3130型自动测序仪上进行测序。
利用Clustal X(1.83)对测序结果进行对位排列,并辅以人工校正。计算群体内和群体间的遗传距离,使用Median-joining法构建单倍型之间的网络关系图,另外还利用分子方差分析(AMOVA)来评估群体间遗传变异。以上分析均在 MEGA 3.1、Arlequin 3.11、DnaSP 4.0、Network 4.1.0.7软件上进行。
2 结果与分析
2.1 外部形态分析
置换检验结果显示,采样点之间群体的计数性状和度量性状均存在差异(P<0.05)。在计数性状上没有发现相关性高的群体组别(图2),同时在计量形态上也不存在相关性高的群体组别(图3),表明中日暂定措施水域南部区域及其以南邻近区域采样点间日本鲭样本组别的形态特征差异大,显分散的状态,无法判断种群属性。
2.2 分子遗传学分析
基于Cytb基因序列的遗传距离显示,日本鲭各单倍型间的遗传距离在0.1%~0.5%,平均遗传距离为0.3%,群体间的遗传距离0.1% (表1)。SHAKLEE综合已发表的资料[14],提出鱼类在属、种和种群三级水平上的遗传距离D值分别为0.90、0.30及 0.05的分类依据,并根据BILLINGTON等[15]认为种内遗传分化应该小于10%的标准,因此认为本研究9个点的样本群体间遗传变异较小。
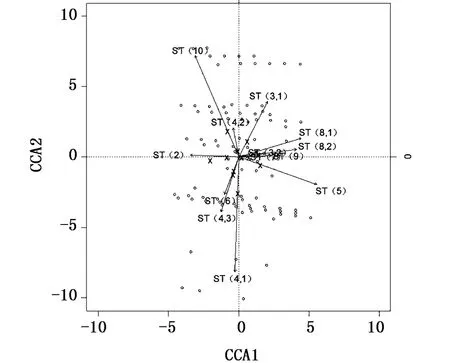
图2 采样点之间样本计数性状的相关性图Fig.2 Correlation of catch count traits between sam p le points
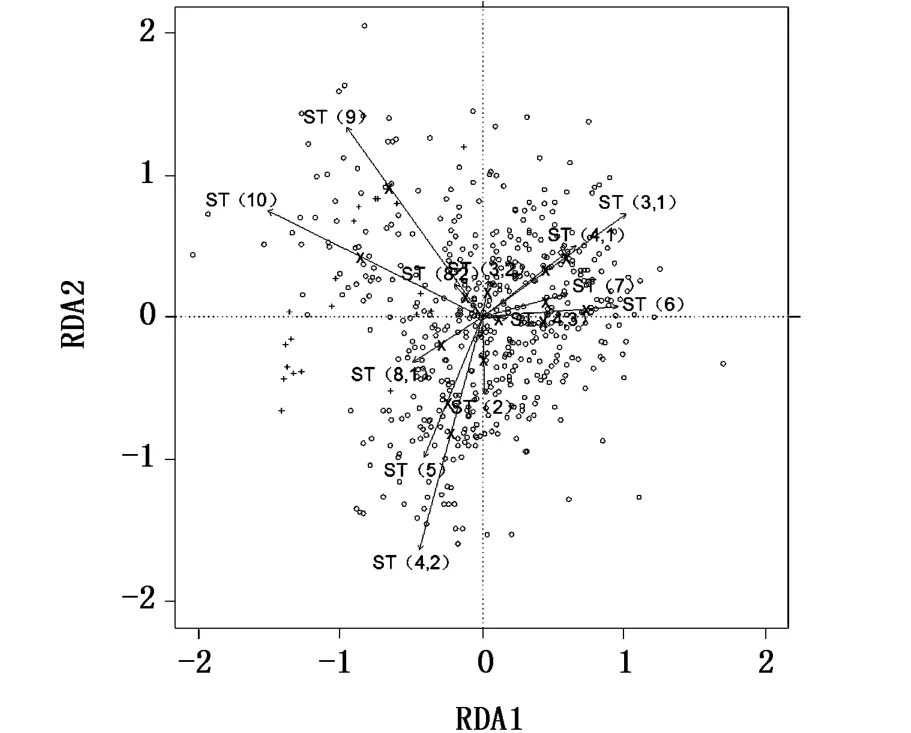
图3 采样点之间样本度量性状的相关性图Fig.3 Correlation of catch measurement characters between samp le points
从日本鲭Ctyb单倍型网状亲缘关系图来研究其单倍型序列遗传结构与样本采集地之间的关系显示(图4),所有的Cytb单倍型成星状分布,H2为中心单倍型,为9群体106个个体共享,单倍型H3与H2相连,为9群体38个个体共享,H1单倍型为5群体共享单倍型,Cytb单倍型序列并没有以采样地形成各自的分支,网络图表明群体间无明显的分化。
对Cytb数据进行AMOVA分析发现,本次分析的9个采样点的日本鲭变异主要来自于群体内,占总变异的97.41%,群体间的变异较小,固定系数为0.025 9(表2),表明群体为随机交配。

图4 基于日本鲭线粒体Cytb基因变异的29个单倍型的网络关系图Fig.4 Network diagram of 29 hap lotypes ofm itochondrial based on Cytb genemutation of Scomber japonicus
综合Cytb序列变异得到的遗传距离、网络关系图及群体AMOVA分析结果,可认为所研究的各采样点的日本鲭无明显遗传分化,初步判定为同一种群。

表1 基于日本鲭线粒体Cytb基因序列的各群体遗传距离Tab.1 Genetic distance of Scomber japonicus based on m itochondrial Cytb gene sequences

表2 基于日本鲭线粒体Cytb基因序列的AMOVATab.2 AMOVA of Scomber japonicus popu lation based on m tDNA Cytb gene sequences
3 讨论
3.1 关于东黄海日本鲭种群的假说
邓景耀等[6]将东黄海的日本鲭划分为东海西部种群、五岛西部种群和闽南-粤东地方种群。然而,丁仁福等[7]将五岛西部群和东海西部群归为东海群系,这一划分与佐美修造[16]将分布于东海不同越冬场日本鲭鱼群作为一个种群-对马暖流群系(中国称谓的东海群系)的观点一致。但是,日本学者[8]将分布于东海中部以北至山阴海域的日本鲭划分为九州西部群系;将在东海南部产卵、夏季到达东海北部及黄海、冬季洄游至东海南部的日本鲭划分为东海西部群系。
日本学者[8]研究表明,分布于日本海西部和东海的日本鲭在计数性状和度量性状上存在地理性变异情况,并将它作为证明多个种群存在的一个证据。刘楚珠等[11]运用形态框架法,发现在东海、黄海的各个渔场捕获的日本鲭繁殖群体都表现出不同的形态特征,使用该方法不能确定东海仅存在一个种群。由此可见,在以往的东黄海日本鲭种群研究中,运用外部形态学鉴定方法的研究结果支持标志放流及渔业调查资料分析存在多个种群的观点。
本研究中形态学方法的结果显示,取自范围相对狭窄的中日暂定措施水域南部区域及其以南相邻区域的采样点间的日本鲭产卵群体表现出各异的形态特征,没有观察到性状类似的群体组别,呈现分散的状态,这与刘楚珠等[11]的研究结果类似。然而,本研究中分子遗传学结果表明研究区域内的日本鲭产卵群体在遗传上没有检测出差别,可判别属于同一种群。日本研究者①福若雅章,由上龙嗣,大下誠二。日中暫定措置水域中心をとする東シナ海のマバ系群に関する共同研究。日本西海区水产研究所,2017年。在2012年2-6月采集了分布于东海南部至对马海峡沿黑潮暖流区洄游的日本鲭繁殖群体的种群判别研究中,利用外部形态方法无法判别种群属性,而利用DNA方法可判别为一个种群,这与本研究的结果是一致的。依据中国和日本在取样区域的部分重迭性和DNA方法的分析结果,推断东海海域的日本鲭为一个种群。
3.2 外部形态特征无法判别东海中南部日本鲭种群的解释
中国学者[17]综合相关研究认为,分布于东海中南部的日本鲭沿西北偏北向进入闽东、浙江近海进行生殖洄游,而日本研究者[18]认为,该海域的日本鲭以东北向作生殖和索饵洄游,可抵达对马海峡,以及产卵场可分为浙江近海、东海外海、九州西部和济州岛近岸。而从本研究2-5月采自东海中南部日本鲭样本性腺发育程度看,1龄及以上个体多数已达3~5期,部分正在产卵或已产完,说明该区域也是产卵场。基于上述日本鲭生殖洄游分布的认知,分布于东海中南部的日本鲭可能是分2股作生殖洄游的,一股朝西北偏北方向沿中国近海生殖洄游;另一股朝东北偏北方向沿东海外海生殖洄游。从渔场和海洋流系配置看,向闽东、浙江近海产卵洄游的群体主要受台湾暖流和闽、浙江沿岸流所控制,而另一股产卵洄游的群体主要受黑潮暖流所控制。因受栖息环境影响,东海分布的日本鲭群体有可能来源于不同发育环境,因此推测不同产卵场的日本鲭幼鱼在外部形态特征上存在差异。
基于东黄海日本鲭越冬结束后,每年春季始于东海中南部生殖洄游的模式[8,17-18],可认为本研究采集于东海中南部海域的日本鲭样本来自于不同产卵场个体的混栖群体,这些样本因在外部形态上存在差异,干扰了形态性状特征矢量值,或许是利用形态特征无法判断种群属性的原因。
综上,基于分子遗传学证据,并且考虑东海中南部渔场的连续性、产卵场位置相近等情况,笔者认为包含中日渔业协定暂定措施水域在内的东海中南部区域的日本鲭群体可视为同一个单元。本研究可作为东海海域日本鲭为一个种群的新证据,佐证丁仁福等[7]和佐美修造[16]提出的东海存在一个种群的观点。
此外,应研究东海中南部日本鲭向近海或向外海生殖洄游时其数量多寡的决定因素及量化模型和鱼卵输送模式,并结合中日围网渔业在东海中南部捕捞日本鲭渔场差异性,对日本鲭资源可分为中日2个独立的评估管理单元,或许更适合于东海中南部日本鲭渔业管理现实。
猜你喜欢
海洋石油(2021年3期)2021-11-05
华人时刊(2020年21期)2021-01-14
高中生之友(中旬刊)(2019年9期)2019-10-23
中华诗词(2019年2期)2019-08-27
赤峰学院学报·哲学社会科学版(2018年9期)2018-12-10
现代农业科技(2016年22期)2017-03-24
诗潮(2014年7期)2014-02-28
环球时报(2009-02-17)2009-02-17
军事历史(1993年3期)1993-08-21
