
基于16S rRNA和COI基因序列的柔鱼亚科系统进化关系探讨
2018-10-26 08:06任桂静唐峰华马春艳张凤英马凌波
海洋渔业 2018年5期
任桂静,唐峰华,马春艳,张凤英,马凌波
(中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,上海 200090)
柔鱼科(Ommastrephidae)隶属于软体动物门(Mollusca)头 足 纲 (Cephalopoda)枪 形 目(Teuthoidea),一般根据形态特征划分为柔鱼亚科(Ommastrephinae)、褶柔鱼亚科(Todarodinae)、滑柔鱼亚科(Illicinae),共计11属23种,大多为大洋性种类,十分强壮,游泳迅速[1]。柔鱼亚科主要分类特征为漏斗浅穴内生纵褶,两侧具边囊,具皮下、内脏和眼球发光器,一般认为柔鱼亚科是柔鱼科进化较晚的一个类群,包括6属9种(图 1,图 2,表 1)[1-2]。随着我国远洋渔业不断发展,柔鱼等重要经济种类的生物学特征、资源动态、渔场环境等已经积累了较多的研究资料[3-6]。对柔鱼亚科系统进化关系的研究主要以形态学为主[2,7-10]。
目前DNA条形码技术已经成功地应用于头足类研究的许多领域,包括种群结构分析、物种鉴定、新种的发现、系统进化和生物多样性研究。SALES等[11]利用16S和COI基因条码,结合一个核基因标记(Rhodopsin基因),发现枪乌贼科的普氏枪乌贼(Doryteuthis plei)、皮氏枪乌贼(Doryteuthis pealei)、杜 氏 枪 乌 贼 (Uroteuthis duvauceli)和莱氏拟乌贼(Sepioteuthis lessoniana)均可能存在隐蔽种。MERIAM等[12]利用COI序列研究了乌贼(S.officinalis)的群体遗传多样性和种群结构。WAKABAYASHI等[13]利用16S和COI基因条码片段探讨了柔鱼科的系统进化关系。YOSHIDA等[14]利用线粒体12S、16S和 COI基因片段研究了乌贼科的系统发生关系。
近年来,贝叶斯联合模型分析多基因片段的方法广泛应用于系统发育研究中[15-16],联合多基因片段分析能有效地增加系统发生分析的信号强度和置信度,并为不同进化速率的片段选择最适进化模型和分组方法。运用贝叶斯法(BI)构建分子系统树,通过“联合”途径获得的结果更可能接近正确的物种树[17-18]。本研究拟利用线粒体COI和16S rRNA基因条形码技术,选取柔鱼亚科7个种(图2)[19],以太平洋褶柔鱼和滑柔鱼为外群,构建贝叶斯联合系统发育树,并结合形态分类特征,探讨柔鱼亚科各种的分类地位和系统发育关系,其研究结果可为柔鱼等资源的保护和利用提供分子水平资料。
1 材料与方法

图1 柔鱼亚科各属系统发生关系树(仿 ROELEVELD[2])Fig.1 Phylogenetic tree of the Ommastrephinae(Figure based on ROELEVELD[2])
1.1 实验材料
本实验所用柔鱼亚科样品共8尾,3尾柔鱼样品于2013年9月中旬远洋捕捞船采自北太平洋区域(42°56′N、158°45′E),2尾茎柔鱼样品于2013年9月中旬远洋捕捞船采自秘鲁外海,1尾鸢乌贼样品于2013年3月上旬采自我国南海,2尾发光柔鱼(Eucleoteuthis luminosa)样品于2015年8月底远洋捕捞船采自千岛群岛附近(45°N、155°E)。以柔鱼科另外两个亚科的代表种—1尾太平洋褶柔鱼(Todarodes pacificus)(褶柔鱼亚科,2015年12月底采自日本海)、1尾阿根廷滑柔鱼(Illex argentinus)(滑柔鱼亚科,2015年7月底采自阿根廷近海)作为外群(表1)。所有样品运回实验室后取肌肉组织用95%乙醇固定,在-20℃条件下保存备用。从GenBank上取得玻璃乌贼(Hyaloteuthis pelagica)、鸟柔鱼、大西洋鸟柔鱼(Ornithoteuthis antillarum)的序列参与分析。

图2 本研究涉及的柔鱼亚科7个种Fig.2 7 species of Ommastrephinae in this study

表1 本研究所用样品信息Tab.1 Information of species used in this study
1.2 基因组DNA提取、片段序列测定
取柔鱼肌肉100 mg左右,用海洋动物组织DNA提取试剂盒提取基因组DNA,-20℃下保存备用。
用于扩增 COI基因的引物为 LCO1490:5’-GGT CAA CAA ATC ATA AAG ATA TTG G-3’和HCO2198:5’-TAA ACT TCA GGG TGA CCA AAA AAT CA-3’[20];用于扩增 16S rRNA基因的引物为16Sar:5’-CGCCTG TTTAAC AAA AAC AT-3’和 16Sbr:5’-CCG GTC TGA ACT CAG ATC ACG T-3’[21]。PCR反应体系总体积为 25μL,其中:10×buffer 2.5μL,MgCl21.5 mmol·L-1dNTP 200μmol·L-1引物各 0.2μmol·L-1,Taq酶1.25 U,模板 DNA 1μL,加灭菌蒸馏水至25μL。PCR反应在Eppendorf Mastercycler 5333扩增仪上进行。反应条件为:94℃预变性3 min,然后进行35个循环,每个循环包括94℃45 s,52℃退火45 s,72℃ 1 min,最后72℃延伸10 min。
以上反应均用阴性对照来检查是否有DNA污染。取1.5μL扩增产物用1.5%琼脂糖凝胶电泳,EB染色,凝胶成像系统检测。扩增产物送上海杰李生物科技有限公司测序。
1.3 数据分析
应用 Dnastar软件包 (DNASATR,Inc.,Madison,USA)对所有序列进行编辑和排序校对。再应用Clustal X1.83对得到的序列进行序列比对。各种序列的多态性使用Arlequin(Ver.3.11;Excoffier等[22])软件进行分析。在 AIC(Akaike Information Criterion)标准下,使用MrModeltest 2.0选择 DNA序列最佳替换模型[23],运用 MrBayes v 3.1.2中的贝叶斯演绎法(Bayesian Inference,BI)构建系统发育树[24]。采用 MCMC(Markov Chain Monte Carlo)算法,以随机树为起始树,运行100000代,每100代取样1次,开始的250个样本作为老化样本(burn-in samples)舍弃。采用分划同质性检验(partition homogeneity test)评价线粒体COI和16S rRNA基因片段联合分析的可靠性[25]。运用MrModeltest 2.0软件分别对以上2个基因片段及COI编码片段的第1、2、3密码子进行分析,获得最适合本研究序列矩阵的核苷酸替代模型及相关参数,选择合适的分组方法,联合2个基因片段构建系统发育树。
2 结果与分析
2.1 各基因片段独立分析
获得了柔鱼亚科6个种的部分COI和16S rRNA基因片段,下载GenBank数据库中的玻璃乌贼、鸟柔鱼、大西洋鸟柔鱼的相关序列,分析了9个种的COI和16S rRNA 2个基因片段的部分序列。COI基因共得到612 bp的序列,检测到206个多态位点,占总序列的33.7%,简约信息位点173个,没有出现碱基的插入缺失,转换颠换比(TS/TV)为2.3;在16S rRNA基因459 bp的基因位点上,发现71个多态位点,占总序列的15.5%,简约信息位点50个,插入缺失8个位点,转换颠换比为2.1。2个片段的转换颠换比都大于 2,说明都尚未饱和[26]。
以阿根廷滑柔鱼、太平洋褶柔鱼为外群,运用最适模型构建贝叶斯系统发育树。基于COI基因片段序列构建的系统树拓扑结构,运用了最适应模型HKY+I+G。结果显示发光柔鱼先依次与鸢乌贼、玻璃乌贼、茎柔鱼聚为一大支,然后再与柔鱼聚合,而鸟柔鱼属两个种(鸟柔鱼和大西洋鸟柔鱼)形成一个单系群(monophyly)且与以上各种分化较大,外群的两个种显示了与以上各种较大的遗传分化,同时该系统树种间各分支都具有较高的后验概率(0.75~1)(图3);基于16S rRNA基因片段序列构建的系统树拓扑结构,运用了最适应模型GIR+I。结果显示了较大的差异,首先柔鱼与鸢乌贼聚为一支,然后依次与玻璃乌贼、茎柔鱼,聚为一大支再与发光柔鱼聚合,且鸟柔鱼属两个种形成一个单系群,表现出与太平洋褶柔鱼关系较近,该系统树各分支的后验概率方面除发光柔鱼外,其余种间后验概率过低(0.21~0.39)拓扑关系可信度较低(图 4)。
2.2 片段联合分析
线粒体COI和16S rRNA基因片段同质性检验的结果显示P值为0.52,说明这2个基因片段可以进行序列联合分析。COI基因为编码基因片段,其第1、2密码子高度保守,而第3密码子容易发生碱基替换,因此大部分系统发生信息存在于第3密码子。将编码片段COI的第1、2和3密码子分别运用MrModeltest2.0软件分析,获得最适合本研究序列矩阵的核苷酸替代模型及相关参数(表2)。因此COI第1、2密码子序列为一组;第3密码子序列为一组;16S rRNA序列单独为一组。根据分组方法和不同分区的最适进化模型,构建系统发育树(图5)。序列联合分析系统发育树显示,其系统结构为发光柔鱼依次与鸢乌贼、玻璃乌贼、茎柔鱼聚为一大支,然后与柔鱼聚合,鸟柔鱼属两个种(鸟柔鱼和大西洋鸟柔鱼)两个种形成一个单系群且与以上各种分化较大,外群的两个种显示了与本研究各种较大的遗传分化。联合分析得到的系统树上各分支后验概率很高,都接近于1(图5)。

图3 基于COI基因片段序列构建的柔鱼亚科贝叶斯系统树Fig.3 Bayes phylogenetic tree of Ommastrephinae based on partial COI gene

图4 基于16S rRNA基因片段序列构建的柔鱼亚科贝叶斯系统树Fig.4 Bayes phylogenetic tree of Ommastrephinae based on partial 16S rRNA gene
3 讨论
位于线粒体基因的16S rRNA和COI基因条码是海洋生物分子系统研究中常用的分子标记,本研究基于这两个基因片段分别构建了柔鱼亚科系统发生树。研究结果的差异主要集中在两方面:1)在鸟柔鱼属分类地位问题上,基于16S rRNA基因片段的系统树显示鸟柔鱼属两个种,脱离了柔鱼亚科范围,表现出与太平洋褶柔鱼关系较近,这种结果与 WAKABAYSHI等[13]研究结果相同,但基于COI基因片段的系统树却显示鸟柔鱼属两个种仍然与柔鱼亚科其它种聚为一大支,这两种结果都具有很高的后验概率。2)在除鸟柔鱼属外,柔鱼亚科内部系统关系问题上,基于COI基因片段的系统树各分支具有较高的后验概率(0.72~1),具有较高的可信度,拓扑结构显示柔鱼和茎柔鱼亲缘关系较近,且为分化较早的种类,而发光柔鱼、鸢乌贼和玻璃乌贼亲缘关系较近,为柔鱼亚科分化较晚的种类,而基于16S rRNA基因片段的系统树各分支的后验概率方面除发光柔鱼(值为1)外,其余种间后验概率过低(0.21~0.39),拓扑关系可信度较低。线粒体16S rRNA基因因其较为保守而被较多地用于种以上阶元的系统发育研究[27-29],而本研究结果显示其在柔鱼亚科属间的分析可能存在一定的局限。WAKABAYSHI等[13]基于16S rRNA基因片段运用3种方法构建的系统树柔鱼亚科内部同样支持率过低,证实了以上观点。
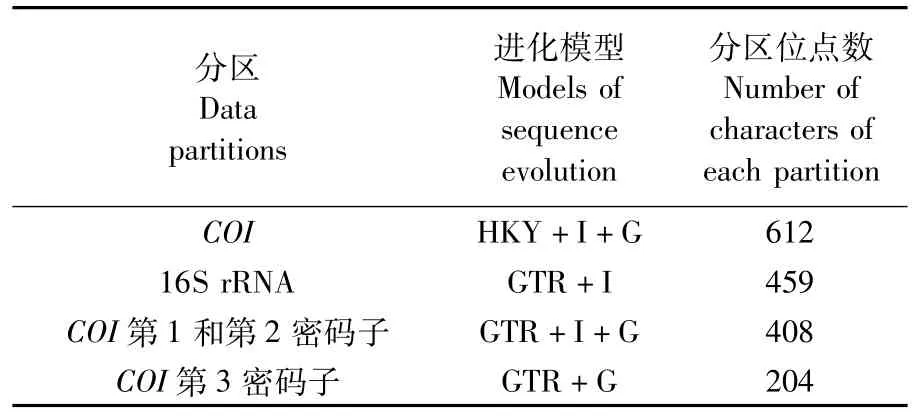
表2 不同分区及其最适进化模型和所包含位点数Tab.2 Data partitions,estimated models of sequence evolution and total number of characters of each partition
由COI片段独立分析得到的结果与联合两个基因条码构建的系统发育树拓扑结果一致,可信度均接近100%。因此我们以此分析结果讨论。鸟柔鱼属在以往大部分研究中因其出现内脏发光器和漏斗浅穴边囊等特征,被归入柔鱼亚科[2,29-31],本研究结果支持以上观点,将鸟柔鱼属仍然归入柔鱼亚科,但属于柔鱼亚科中最原始种类。JEREB等[7]研究认为柔鱼科种类在进化过程中经历了陆架坡、浅洋性、完全大洋性三个阶段。鸟柔鱼属种类属浅洋性种类,本研究结果显示鸟柔鱼属可能是褶柔鱼亚科向柔鱼亚科进化过程中的过渡物种。

图5 基于COI和16S rRNA基因片段序列构建的柔鱼亚科贝叶斯系统树Fig.5 Bayes phylogenefic tree of Ommastrephinae based on partial COI and 16S rRNA gene
在以往的形态学研究中,如ROELEVELD[2]、ZUEV等[8]认为玻璃乌贼属、发光乌贼属亲缘关系较近,最显著的形态特征是外套膜上都具有特化的大型发光器(玻璃乌贼属具圆斑点,发光乌贼属具条状)(图2),而鸢乌贼与茎柔鱼属亲缘关系近。但是,基于分子数据的研究结果发现玻璃乌贼、发光乌贼和鸢乌贼亲缘关系较近。通过仔细研究鸢乌贼属形态特征发现其外套膜后背前部一大块皮下发光器的集聚,形成一个大型发光器(图2)。整个柔鱼科种类中只有玻璃乌贼、发光乌贼和鸢乌贼3属均有外套膜大型发光器[7],可见本研究结果有一定的形态学依据。
本研究中柔鱼与其它柔鱼亚科种类存在较大的遗传距离(枝长较长),可能提示有其独特的进化路径。从发光器这一特征来看,柔鱼属种类只有外套膜皮下发光器,这与柔鱼亚科其它种类具有较大差别(其它种类包括最原始的鸟柔鱼都具有内脏和眼球发光器),ROELEVELD[2]研究认为这是柔鱼亚科某祖先种在进化过程中丢失了内脏和眼球发光器,产生了外套膜皮下发光器,进而形成了柔鱼属种类。本研究显示柔鱼与其它柔鱼亚科种类存在较大的遗传距离,可能预示着柔鱼亚科种类一个新的进化方向。
综上所述,玻璃乌贼、发光乌贼和鸢乌贼亲缘关系较近,是柔鱼亚科中分化较晚的种类,其次为茎柔鱼、柔鱼,鸟柔鱼属两个种为柔鱼亚科中最原始种类,且为褶柔鱼亚科向柔鱼亚科进化过程中的过渡物种。本研究同时显示了各类发光器在柔鱼亚科系统发育研究中具有非常重要的进化意义。但由于本研究样品种类尚不完善,研究所用线粒体DNA是母系遗传,仅能反映母本的遗传特征,并且其各基因片段进化速率差异较大。今后有必要增加核基因标记的研究,并结合更多形态学特征,进一步完善柔鱼亚科分类地位和系统发育关系。
猜你喜欢
山西大学学报(自然科学版)(2021年4期)2021-08-31
法律方法(2021年4期)2021-03-16
法律方法(2021年4期)2021-03-16
军事文摘(2020年14期)2020-12-17
统计与决策(2019年6期)2019-04-22
智富时代(2018年11期)2018-01-15
智富时代(2018年11期)2018-01-15
雷达学报(2017年6期)2017-03-26
家教世界·创新阅读(2016年12期)2017-01-09
阅读与作文(小学高年级版)(2016年1期)2016-03-04
