
贝类闭壳肌-贝壳连接界面的分子组成及连接机制
2018-09-19 08:38姜雨婷
浙江海洋大学学报(自然科学版) 2018年4期
廖 智,孙 琦,姜雨婷
(浙江海洋大学海洋科学与技术学院,浙江舟山 316022)
贝壳是多数软体动物用以保护其内部软组织的一层硬壳,其主要成分是碳酸钙,占贝壳总质量95%以上。作为一种高度结构化的,具有优异力学性能的天然纳米复合材料,贝壳相关研究不仅是仿生学及生物材料学研究的重要领域,同时,在生物医学工程以及生物材料工程方面也显示出广阔的应用前景[1-4]。目前,针对贝壳的研究主要包括贝壳微观结构的类型、组装、碳酸钙晶体结构及形成过程分析,贝壳中无机质和有机质成分组成以及组装特征分析,贝壳中蛋白质(即贝壳基质蛋白)的分子组成、结构以及对碳酸钙晶体形成的影响分析等。此外,贝类的肌肉与贝壳之间的连接界面,如蜗牛的中柱肌-贝壳,双壳贝类的后闭壳肌-贝壳连接界面等,也引起人们的关注。贝类中柱肌或闭壳肌均属肌肉组织,而贝壳是由碳酸钙晶体构成的非细胞组织,两者之间的连接不仅涉及生物大分子之间的相互作用,还涉及有机物-无机物之间的结合,其本质属于生物粘附研究范畴。考虑到目前肌肉与生物硬组织之间的生物粘附是生物医学工程和生物材料学的重要研究内容,也是组织工程材料的研发,植入,修复和替代等研究的基础。因此,贝类的闭壳肌与贝壳的连接界面研究一方面可以为组织工程材料在人体中的移植修复等过程中,构建材料与受体组织间的粘附提供很好的仿生学启示;另一方面,该界面研究所涉及的,参与连接的关键蛋白分子可作为先导分子,在新型人工肌腱材料的研发中具有重要意义。
1 贝壳与贝壳基质蛋白
对贝壳的研究起始于对贝壳结构的微观观察和力学分析。在早期研究中,人们已注意到贝壳具有极为复杂的微观结构。随着技术进步,近20年来,人们开始关注贝壳中各种有机质成分,特别是蛋白质分子在贝壳中的组成、分布以及对贝壳形成的影响,相关研究逐年增多。以mollusc shell作为关键词搜索Pubmed数据库(https://www.ncbi.nlm.nih.gov/pubmed/?term=mollusc+shell),可见1970-2017年,贝壳相关研究的年发表文章数量呈明显上升趋势(图1)。上述研究为了解贝壳肌肉-贝壳连接界面奠定了基础。而针对软体动物肌肉-贝壳连接界面的研究则在上世纪七十年代有少量结构分析报道,之后长期处于停滞状态。随着对贝壳结构以及贝壳基质蛋白组成的深入了解,这一方面的研究在近几年才逐渐受到重视,但总体而言,相关研究仍然极少。由于贝类肌肉-贝壳的连接界面的研究是建立在贝壳自身结构及贝壳基质蛋白的结构与功能研究基础之上,因而本文首先关注贝壳与贝壳基质蛋白的研究进展。

图1 1970-2017年,Pubmed数据库收录的贝类贝壳相关研究文章的发表数量Fig.1 The number of published papers in the field of mollusc shell at 1970-2017 from Pubmed database
贝壳是由碳酸钙晶体经过层层组装而成。贝壳中的碳酸钙晶体其主要晶型为方解石(Calcite)和文石(Aragonite)两种物相结构[5-7]。根据碳酸钙晶型和形貌的区别,可将贝壳结构分为不同的微观结构层次,常见的有三种,即珍珠质层(Nacre),棱柱层(Prismatic layer)和斜交叉层(Obliquely Crossed Crystals)[8-9]。上述分层仅是从贝壳的整体结构而言所做的一个简单分层,事实上,不同贝类的贝壳结构,不论是整体外形还是微观结构组成方面,均表现出明显的结构多样性特征[10-11]。根据CARTER[11]对不同贝类贝壳的扫描电子显微镜观察结果,报道了至少七组不同类型的微结构,包括棱柱组(Prismatic)、球状组(Spherulitic)、板状组(Laminar)、交叉组(Crossed)、均质组(Homogeneous)、孤立针状组(Isolated spicules and spikes)以及孤立晶体组(Isolated crystal morphotypes)。不同贝类的贝壳中,其微观结构的组装形式和种类不一样,但通常包括上述2~7种结构层次。不同的微观结构层次往往也具有不同的力学性能,目前发现贝壳中,参与和肌肉连接的结构层次主要是肌棱柱层(myostracum),属于棱柱组类型。
贝壳所具备的优异力学性能不仅来自于碳酸钙晶体在纳米层次的结构组装,也来自于贝壳中各种有机质成分的辅助[12]。有机质是贝壳的重要组成成分,尽管其含量较低,不到贝壳总质量的5%,但对贝壳的形成以及贝壳的力学性能起到了至关重要的作用。特别是其中的蛋白质,贝壳基质蛋白(Shell Matrix Protein,SMP),直接影响了贝壳碳酸钙晶体的成核以及定向生长等过程[12-15];而贝壳中的其他有机物,如几丁质等成分,在贝壳的贝壳无机相与有机相的分子连接以及构建贝壳框架结构中发挥着关键作用[16-17]。在贝壳有机物的指导和调控下,最终将碳酸钙晶体沉积形成具有不同微观结构层次的贝壳,这一过程被称之为生物矿化(Biomineralization),这也是一个无机相-有机相不同分子的相互识别和相互作用的过程[18]。同时,贝壳基质蛋白中,也必然有一部分蛋白参与了贝壳与肌肉组织之间的连接,因而贝壳基质蛋白的研究为从中筛选参与肌肉-贝壳连接的关键蛋白奠定了基础。
目前已报道的贝壳基质蛋白种类众多,目前在美国国立医学图书馆的蛋白质数据库(National Center for Biotechnology Information Search database)中,以“molluscs shell matrix protein”作为关键词可搜索到的蛋白质种类达到785种,可归属于37个蛋白质家族(https://www.ncbi.nlm.nih.gov/ipg?term=molluscs%20shell%20matrix%20protein)。其物种来源涉及142种软体动物,绝大多数来自双壳纲Bivalves,少数来自腹足纲Gastropods和头足纲Cephalopods。其分子大小从95个氨基酸残基(数据库编号:AAQ08227.1)到1 912个氨基酸残基(数据库编号:JAP63242.1)不等。就单独的贝类贝壳而言,一种贝类贝壳中所包含的贝壳基质蛋白种类推测在200种以上[19],但在实际研究时,由于贝壳基质蛋白的含量极低,且通常水溶性极差,因而即使采用蛋白质组学策略,往往也只能鉴定到其中数十种丰度较高的基质蛋白。例如,UPADHYAY,et al[20]从牡蛎Crassostrea hongkongensis和C.angulata的贝壳中分别鉴定到42和37种贝壳基质蛋白;ARIVALAGAN,et al[21]从蛤 Mya truncata的贝壳中鉴定到 67种贝壳基质蛋白;LIU Chuang,et al[22]从珠母贝Pinctada fucata的贝壳中鉴定到72种贝壳基质蛋白;笔者也分别从地中海贻贝Mytilus galloprovincialis和厚壳贻贝M.coruscus贝壳中分别鉴定到113和63种贝壳基质蛋白等[23-24]。已报道的贝壳基质蛋白往往具有特定的结构域或特殊的氨基酸组成,且在贝壳不同微观结构层次中也具有不同的分布特征[25]。因此可以根据贝壳基质蛋白在贝壳不同层次或者不同晶型碳酸钙中的分布大致可将贝壳基质蛋白分为珍珠质层特有基质蛋白,棱柱层特有基质蛋白等,或方解石层特有基质蛋白,文石层特有基质蛋白等[26]。由于相关综述较多,因此笔者在此不做赘述。但值得注意的是,由于参与贝类闭壳肌-贝壳连接的主要结构层次是属于棱柱层的肌棱柱层,因而该层所特有的贝壳基质蛋白很有可能参与了该界面的连接。
上述研究不仅使人们了解贝壳自身的结构特征及贝壳基质蛋白的种类和分布特征,同时也发现参与肌肉连接的贝壳部位与非肌肉连接的贝壳部位在微观结构和蛋白质分子组成方面具有明显差异,这为贝类肌肉-贝壳连接界面的相关研究起到了重要的推动作用。
2 贝类肌肉-贝壳连接界面的连接模型
贝壳中,特别是双壳贝类的后闭壳肌-贝壳连接界面(adductor-muscle-shell attachment,AMS)不仅涉及生物大分子之间的相互作用,还涉及有机物-无机物之间的粘附。目前在人造骨骼移植以及人工肌腱材料的临床应用中,均面临人造骨骼和肌肉间不易产生粘连,粘附力不足易断裂以及生物相容性差等问题[27]。而贝类的闭壳肌与贝壳的连接界面涉及到两种不同的组织体系(肌肉和贝壳)间的连接,因而该连接界面的研究对相关生物医学和材料学研究具有很好的启示意义。因此,对于贝类肌肉与贝壳之间的连接,人们首先关注的是其力学性能以及该界面的微观结构。双壳贝类闭壳肌通过紧密结合贝壳,可将两片贝壳紧紧的拉在一起(图2)。对双壳贝类而言,闭壳肌收缩时产生的作用力极强,GALTSOFF[28]曾报道,牡蛎C.virginica的贝壳闭合后,需20 kg的拉力方可将其拉开;而在强行拉开牡蛎闭合的双壳过程中,断裂部位通常发生在闭壳肌内部,而闭壳肌与贝壳的连接界面仍然保持完整。上述研究一方面表明闭壳肌本身具有较强的收缩力,另一方面也表明贝类闭壳肌与贝壳之间的连接极为紧密,具有很高的连接强度。考虑到闭壳肌主要成分为肌纤维,而贝壳主要成分为碳酸钙晶体,这两者之间的连接作为一种典型的无机相-有机相连接模式,人们首先关注的是其连接结构。
目前,对于贝类肌肉-贝壳连接界面微观结构的研究早期主要集中在鹦鹉螺以及腹足类的中柱肌-螺壳的连接界面,例如 ISAJI,et al[29]对于鹦鹉螺以及 TOMPA,et al[30]对陆生蜗牛的中柱肌-壳连接界面的微观结构研究;在上述研究中,研究者提出了一个“上皮(肌腱)细胞-半透明膜-贝壳”(epithelium-semitransparent membrane-shell junction)的连接模型(图3)。肌肉细胞通过肌腱细胞(tendon cell)实现与壳之间的连接。同时,上述研究还观察到肌肉中有少量纤维组织深入到壳层内部,从而有利于强化这种连接。
虽然该模型从细胞层面对肌肉-贝壳连接界面的结构特征进行了描述,但是该模型是建立在鹦鹉螺以及蜗牛的中柱肌与壳的连接方式基础上,而中柱肌-壳的连接强度远远低于双壳贝类的闭壳肌-贝壳连接[30];另一方面,该模型侧重于细胞及组织水平构建一个连接模型,而对于在这一连接界面中有哪些分子起到了关键作用以及相关连接的分子机理并未涉及。对双壳贝类的闭壳肌-贝壳连接界面研究起始于对连接界面的微观结构观察,例如早期NAKAHARA,et al[31]利用电镜观察了贝类Pinctuda rudiatu闭壳肌-贝壳连接界面,SONG Yingfei,et al[32]对地中海贻贝的后闭壳肌-贝壳的连接界面的电镜观察,以及近期ZHU Yaoyao,et al[33]对三种双壳贝后闭壳肌痕的微观结构观察。上述研究报道了该界面的微观结构,特别是贝壳部分的碳酸钙晶体结构,而并未涉及肌肉-贝壳的分子组成以及连接机理。上述研究虽然提供了一个贝类-肌肉连接的模型,但相关分子机制仍依赖于贝壳类各种基质蛋白的研究,特别是需要通过蛋白质组学手段首先判断贝壳与肌肉连接的部位有哪些蛋白质分子的存在,且这些蛋白不涉及贝壳自身晶体结构的形成,而是涉及两种界面的连接或者粘附,这方面的工作仍然任重而道远。
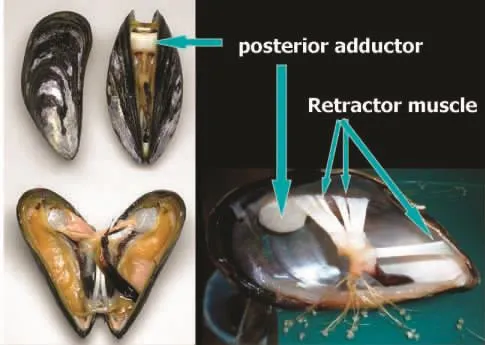
图2 厚壳贻贝后闭壳肌(posterior adductor)以及伸(缩)足肌群(retractor muscle与贝壳内层连接部位(图片摘自笔者未发表数据)Fig.2 The posterior adductor between shells and the retractor muscles between mussel foot and shell in M.coruscus (from unpublished data)
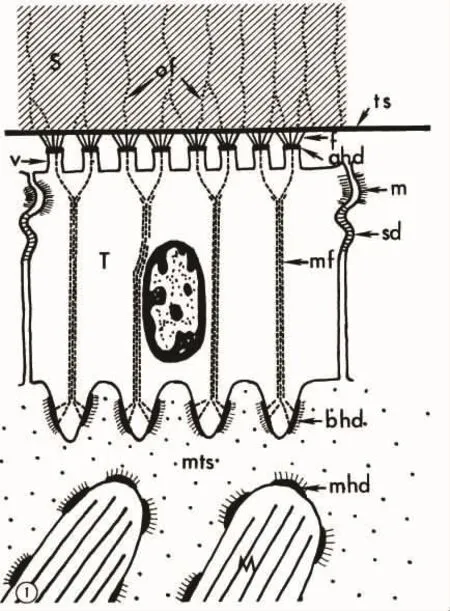
图3 蜗牛(Gastropod)中柱肌与壳连接的模式图(摘自TOMPA,et al[30])Fig.3 Diagrammatic representation of the gastropod muscle attachment to the shell (from TOMPA,et al[30])
3 双壳贝类后闭壳肌痕的微观结构和蛋白质分子组成
在双壳贝类中,闭壳肌与贝壳连接的部位被称为后闭壳肌痕(Adductor Muscle Scar,AMS)[34-35]。在该部位,后闭壳肌肌纤维直接连接于贝壳内侧,其连接部位的主要贝壳结构层次为肌棱柱层(Myostracum)。肌棱柱层为单层柱状结构,在外型上与珍珠质层和纤维棱柱层差异较大,在贝壳中所占比例较低且往往包埋在贝壳内部(图4),但在肌肉-贝壳连接界面,即后闭壳肌痕部位,该层暴露于贝壳内侧表面并直接与后闭壳肌连接,这意味着肌棱柱层在贝壳与肌肉的连接中发挥着重要作用,也必然存在某种分子机制将肌棱柱层与后闭壳肌两种完全不同的物相体系紧密结合在一起。肌棱柱层碳酸钙晶型通常为文石型[36-37]或者文石型和方解石型混合[32]。GAO Peng,et al[23]的研究表明,地中海贻贝贝壳的肌棱柱层中,含有34种肌棱柱层特有的基质蛋白,其中,包括Calponin类蛋白,Transgelin类蛋白,含THY结构域(thymosin beta actinbinding motif)的蛋白等。上述蛋白序列中含有Calponin,Calponin-homology以及THY等具有肌动蛋白结合功能的结构域。推测肌棱柱层中该类型蛋白可能参与了与闭壳肌中肌动蛋白的结合,从而使得肌棱柱层与闭壳肌组织紧密的连接在一起,最终介导了贝壳的闭合[38]。此外,在地中海贻贝以及厚壳贻贝肌棱柱层中还发现丰富的胶原类蛋白[23-24];而此前已知胶原蛋白酶处理可以有效地降低牡蛎C.virginica后闭壳肌与贝壳之间的结合力[28]。上述研究表明,胶原类蛋白可能也在贝壳-肌肉的连接中发挥着重要作用。此外,ZHAO Che,et al[39]的研究表明,在虾夷扇贝Patinopecten yessoensis闭壳肌-贝壳连接部位的表面结构中,存在直径约为7 μm的孔洞结构(Pit structure),并推测该孔洞可能是胶原蛋白在闭壳肌-贝壳连接界面的贯穿部位。对贻贝的后闭壳肌痕表面结构研究也有类似发现[40-41]。同时,在研究中,还发现地中海贻贝的后闭壳肌与肌棱柱层表面之间存在一层有机质膜,且发现肌肉组织与该膜之间存在纤维状物质的连接[41](图5),同时,SONG,et al[32]发现在贻贝贝壳内部也存在纤维状连结物质。上述研究表明,闭壳肌与贝壳肌棱柱层之间的连接可能不仅有成膜蛋白的参与,也有纤维类蛋白质的参与,由此形成了肌肉-膜-贝壳的连接体系,且该体系有胶原类蛋白贯穿并加强了该连接界面的强度。

图4 几种代表性贝类的肌棱柱层电镜图像Fig.4 The SEM photo of myostracum from some representative bivalves

图5 贝类后闭壳肌痕部位表面结构Fig.5 The SEM photo of AMS surface from some representative bivalves
目前,关于闭壳肌-贝壳连接界面的研究由于涉及生物化学,分子生物学,生物力学,生物物理学等多个学科,综合性较强,研究难度较大,相关研究也较少,目前对该界面连接的分子机制也多数处于推测状态,但随着研究的深入,人们必将深入了解该连接界面的分子机制并对仿生学和生物材料学研究产生深远影响。
猜你喜欢
石油沥青(2021年5期)2021-12-02
红领巾·探索(2021年8期)2021-08-26
石材(2020年12期)2020-12-31
考试与评价·高二版(2020年6期)2020-09-10
无机盐工业(2020年4期)2020-04-15
食品与生活(2019年8期)2019-10-30
建材发展导向(2019年5期)2019-09-09
新高考·高一数学(2019年3期)2019-09-07
食品与生活(2017年11期)2017-12-18
高中生学习·高二版(2016年8期)2016-05-14
