
原核表达并纯化沙眼衣原体(Ct)巨噬细胞感染增强因子MIP的免疫活性分析
2018-09-18 03:33史桂桃陆春雪
中国中西医结合皮肤性病学杂志 2018年4期
史桂桃,陆春雪
(1.内蒙古医科大学附属医院,内蒙古呼和浩特,010050;2.南华大学医学院病原生物学研究所,湖南衡阳 421001)
沙眼衣原体(Chlamydia trachomatis,Ct)是一类专性细胞内寄生的原核细胞型微生物,包括A、B、C、D等19种血清型,其中D-K型可引起人类泌尿生殖道感染。Ct是细菌性性传播疾病最常见的病原体,感染者可出现盆腔炎、不孕不育等严重并发症[1]。目前,Ct感染临床治疗最棘手的问题是绝大部分感染者初期症状不典型,一般不会寻求系统的治疗,从而使下生殖道入侵的病原体得以逐渐上行感染至上生殖道;此外,Ct自然感染后免疫力不持久及性伴侣的隐性感染均导致Ct感染人群不断扩大,显然研制出新型有效的Ct疫苗是预防控制Ct疾病的关键[2]。
脂蛋白是细菌胞壁的结构成份,也是分枝杆菌和螺旋体等无脂多糖(LPS)细菌的重要致病物质及疫苗研究靶标[3],衣原体细胞壁LPS缺乏内毒素活性,提示其脂蛋白也可能具有重要作用。巨噬细胞感染增强蛋白(Microphage infectivity potentiator,MIP)是衣原体胞壁重要的脂蛋白成份,可体外刺激巨噬细胞产生并释放前炎症因子。本课题组前期采用自行制备的MIP单克隆抗体证实,MIP蛋白结构保守,在沙眼衣原体各血清型广泛分布,且MIP单克隆抗体能有效中和Ct的感染性,可能是重要的潜在Ct疫苗候选分子[4]。本研究采用基因重组技术将Ct D型MIP编码基因定向克隆到原核表达载体pGEX-6p-l,再将重组质粒转化E.coli XLl-Blue,实现了MIP在E.coli中高效表达,并对表达的蛋白进行纯化及免疫活性分析,为研究MIP亚单位疫苗奠定了基础。
1 材料与方法
1.1 材料
1.1.1 菌株 Ct血清型D标准株为本室保存株[5]。
1.1.2 细胞与动物 HeLa-229细胞株购自美国标准菌株保存中心(ATCC,cat#CCL2);Balb/c小鼠由南华大学实验动物部提供。
1.1.3 主要试剂 Pfx DNA聚合酶、BamH I、Not I限制性内切酶、DNA及蛋白相对分子质量标准、Glutathione Sepharose 4B Beads等为Invitrogen公司产品,Cy3 标记的羊抗鼠 IgG(H+L)、IgGl、IgG2a、IgG2b和IgG3购自Jackson ImmunoResearch公司,PreScission Protease(GST融合蛋白剪切酶)为GE Healthcare公司产品,QuickClean PCR纯化试剂盒、质粒提取试剂盒购自美国Qiagen公司。
1.2 方法
1.2.1 MIP基因原核表达载体的构建 从GenBank获取 Ct D型标准株 MIP基因全长序列(GI:15605270),设计并合成一对特异引物,上游、下游引物中分别引入BamHI、NotI限制性酶切位点。引物如下(下划线为酶切位点):5’-CGC-GGATCCATGAAGAATATATTAAGTTGGATG-3’;DNA 为 模板,PCR扩增MIP基因,酶切后插入pGEX-6p-l原核表达载体,转化E.coliXLl-Blue;测序鉴定。
1.2.2 pGEX-6p-l/MIP重组质粒融合蛋白的表达和纯化 将鉴定后的阳性克隆扩大培养,IPTG诱导表达3 h后离心收集菌体,超声破菌上清与Glutathione Sepharose 4B Beads室温结合1 h后,采用GST融合蛋白剪切酶(该酶特异性结合Beads并精确剪切GST标签与目的蛋白间 LeuGluValLeuPheGlnGlyPro基序的Gln-Gly残基)室温作用Beads 2 h,离心收集酶切后的上清液(主要含酶切后的目的蛋白)及Beads洗脱液(非特异性结合的酶切后目的蛋白),最后采用超滤离心管(Millipore,Billerica,MA)离心浓缩,获得纯化的无GST标签纯蛋白。SDS-PAGE电泳观察蛋白表达情况和纯化效果。
1.2.3 动物免疫 雌性Balb/c小鼠共15只,随机分为EB免疫组、MIP蛋白免疫组及PBS对照组,每组 5 只。将 50 μL 1×106IFU 的纯化 EB、30 μg纯化的MIP蛋白或PBS,分别与10 μg CpG混匀后,各加入等量油性弗氏不完全佐剂(IFA),超声乳化后,注射于小鼠两侧臀部。每只小鼠于第0天,14天,28天各免疫1次,共免疫3次。小鼠在每次免疫前及最后1次免疫后2周,尾静脉采血,分离血清,-80℃保存备用。
1.2.4 小鼠特异性抗体及亚型检测 ELISA测定EB免疫组、MIP蛋白免疫组及PBS对照组血清总抗体和IgG亚类。以10 μg/mL EB菌体包被酶标板,4℃过夜,1%BSA室温封闭1 h,加入稀释好的血清,37℃孵育1 h,0.05%PBST洗涤3次后分别加入羊抗鼠总抗体或 IgG1,IgG2a,IgG2b,IgG3 抗体孵育1 h,加入底物ABTS,37℃避光反应15 min后,酶标仪读取405 nm波长的吸光值。
1.2.5 细胞因子检测 末次免疫2周后,处死小鼠,无菌条件下分离脾脏,制备脾细胞悬液;用含10%FCS的RPMI 1640培养液调整细胞数为5×106/mL,加入24孔板,1 mL/孔,每份脾淋巴细胞悬液设3个组,第1组用紫外线灭活的EB(UV-EB)刺激,第2组用10 μg重组的MIP抗原刺激,第3组加入培养基作为对照,37℃,5%CO2的培养箱中刺激培养72 h后收集上清,酶联免疫吸附实验(ELISA)法定量检测Th1型细胞因子IFN-γ的产生水平。
2 结果
2.1 pGEX-6p-l/MIP原核表达质粒的构建 PCR扩增CtD血清型MIP全长基因(编码产物:M1-E243),1%琼脂糖凝胶电泳,扩增片段长度约为729 bp,与预期结果一致,见图1。经BamH I和Not I双酶切重组质粒以及对测序结果的BLAST分析证实克隆质粒序列完全正确。
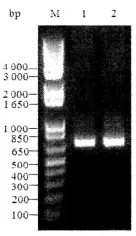
图1 MIP基因的PCR扩增产物电泳图
2.2 pGEX-6p-l/MIP重组质粒融合蛋白的表达与纯化 重组质粒转化的E.coli XLl-Blue,经IPTG诱导表达后获得的表达产物,与Glutathione Sepharose 4B beads结合,得到与Beads结合的GST-MIP融合蛋白复合物,再用特异性的GST融合蛋白剪切酶酶切复合物以去除GST标签,获取游离的MIP蛋白。将Beads-GST-MIP复合物,酶切后上清、Beads洗脱液及酶切后的Beads均用12%SDS-PAGE胶电泳分析,考马斯亮蓝染色结果如图2所示,在相对分子质量约53 KD处可见一清晰条带,与预测的重组MIP融合蛋白相对分子质量一致(GST相对分子质量约26 KD,MIP相对分子质量约27 KD);酶切后无GST标签的纯化蛋白MIP大量存在于酶切上清及洗脱液中,收集酶切后的上清及洗脱液,经超滤离心管浓缩后结果见图3。

图2SDS-PAGE检测蛋白表达和纯化

图3 无GST标签MIP蛋白纯化终产物SDS-PAGE分析
2.3 小鼠血清抗衣原体EB特异性IgG及亚型检测 重组表达的MIP蛋白免疫小鼠后产生的抗体,能有效识别衣原体EB包被的ELISA反应孔,而对照组小鼠血清未检测出特异性的EB抗体;MIP蛋白免疫组小鼠,产生的IgG亚型与EB全菌免疫接种组类似,以IgG2a为主,且IgG2a/IgG1比值大于1,见图 4。

图4 抗血清中IgG滴度及亚型的检测
2.4 MIP免疫可诱导特异性回忆反应 小鼠免疫3次后,取脾细胞,体外用UV-EB、MIP蛋白或无关抗原(细胞培养基)刺激,测定脾细胞上清IFN-γ水平。MIP免疫组小鼠脾细胞受EB和MIP蛋白的刺激,均能产生异性回忆应答,分泌的IFN-γ(Th1型细胞因子)水平明显高于相应的PBS对照组,见图5。但用无关抗原(Medium)刺激时,IFN-γ水平与对照组相比无明显变化。
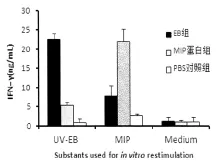
图5 MIP蛋白免疫对细胞因子IFN-γ产生的影响
3 讨论
沙眼衣原体泌尿生殖道感染防治形势日趋严峻,但目前仍缺乏有效疫苗[1]。许多Ct亚单位疫苗,如Ct主要外膜蛋白MOMP[6]、包涵体膜蛋白IncA[7]等被证实有一定的保护作用,但均不完全。这可能因为衣原体血清型别多、候选的抗原分子免疫原性不够强或构象复杂、易变异等因素有关;也可能因为衣原体生活周期复杂,单一抗原难以在Ct吸附、穿入、增殖、释放等各环节发挥作用。鼠衣原体动物实验研究证实采用多种亚单位疫苗联合免疫小鼠产生的保护性效果优于单一疫苗,开发新的亚单位疫苗候选抗原将为联合疫苗的研制提供更多选择。
Ct致病机制不清是制约疫苗成功研制的重要因素之一。Wang等[8]采用体外融合表达的908个Ct开放读码框编码蛋白,与99份Ct泌尿生殖道感染妇女的血清进行反应,发现Ct MIP蛋白能被40%以上感染妇女血清优势识别,是衣原体自然感染过程中优势表达的抗原;Zeng等[9]的动物实验亦证实MIP抗体在90%以上感染小鼠存在,其抗体滴度明显高于MOMP、CPAF和Pgp3等优势抗原[9]。以上的研究均提示MIP蛋白与衣原体的感染致病密切相关,并可能是Ct潜在的新型亚单位疫苗候选分子。
本研究采用基因重组技术成功构建了pGEX-6p-1/MIP重组质粒,并在E.coli中高效表达GSTMIP融合蛋白,经GST融合蛋白剪切酶作用后,获得了高浓度的无标签MIP纯蛋白。纯化的蛋白免疫接种小鼠,可诱导小鼠产生高滴度的针对衣原体EB的特异性抗体,这与笔者前期报道的MIP单克隆抗体具有中和作用的结果一致,说明MIP免疫可提供抗衣原体感染的保护性体液免疫应答;抗体亚型以IgG2a和IgG1为主,二者比值大于1,这种抗体亚型的产生情况提示MIP蛋白免疫可诱导产生Th1型为主的细胞免疫应答;MIP免疫小鼠的脾细胞,在体外接受EB和MIP抗原的刺激,能分泌高水平的IFN-γ,进一步证实Th1型特异性回忆应答的存在。已知Ct为胞内感染菌,抗Ct感染的细胞免疫主要依赖于CD4+Th1型细胞反应,CD4+Th1型细胞及Th1型细胞因子IFN-γ等在清除衣原体感染中起着极其重要的作用[10]。
总之,本实验采用基因工程方法原核表达纯化的MIP蛋白,不含GST标签、且纯度高;接种小鼠后诱导产生的抗体和效应T细胞均能有效识别衣原体菌体EB,发挥免疫保护作用;加之,MIP蛋白结构保守,在Ct各血清型广泛分布,可望成为新型有效的Ct候选亚单位疫苗。
猜你喜欢
动物医学进展(2022年11期)2022-12-28
首都食品与医药(2022年21期)2022-12-06
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
医药前沿(2021年25期)2021-10-16
江西农业学报(2021年4期)2021-04-20
养猪(2020年6期)2021-01-27
家庭科学·新健康(2020年6期)2020-07-06
三农资讯半月报(2020年11期)2020-06-21
浙江畜牧兽医(2019年6期)2019-12-23
