
黄顶菊与3种本地植物竞争对丛枝菌根真菌种类和多样性的影响
2018-09-12 09:44孔令杰韩月龙贾月月张风娟
生物安全学报 2018年3期
孔令杰,柳 旭,韩月龙,杨 康,贾月月,张风娟
河北大学生命科学学院,河北 保定 071002
丛枝菌根(arbuscular mycorrhizal,AM)真菌能与80%以上的陆生植物形成菌根共生联合体( Wardleetal.,2004;Xiaoetal.,2014),是土壤微生物区系中最重要的成员之一(徐黎明,2010)。AM真菌与植物根系形成的共生体是AM真菌与植物之间进行营养交换的场所(Inderjit & Putten,2010)。AM真菌可通过根外菌丝扩大植物根系吸收范围,提高寄主植物对水分和养分的吸收(Fumanaletal.,2006),以促进植物生长。大多数外来入侵植物是菌根植物(姜娜,2013),入侵植物可以直接或间接改变入侵地土壤AM真菌的结构与功能(Hawkesetal.,2006),如紫茎泽兰EupatoriumadenophoraSpreng.入侵后,土壤中AM真菌群落富集了缩球囊霉Septoglomusconstrictum和2种不可培养的球囊霉属的种(任玉晶,2011);加拿大一枝黄花SolidagocandensisL.入侵后,土壤中AM真菌由以摩西球囊霉Funneliformismosseae和幼套球囊霉Glomusetunicatum为优势种转为以地球囊霉G.geosperum和幼套球囊霉为优势种(唐建军等,2009);豚草AmbrosiaartemisiifoliaL.入侵后,土壤AM真菌增加了新的优势种G.iranicum(张玉曼,2015)。所以,研究外来植物入侵对土壤AM真菌结构和多样性的影响有助于揭示其入侵扩散机制。
黄顶菊Flaveriabidentis(L.) Kuntze.是一种入侵我国的外来有害植物(陈冬青,2012; 张天瑞,2011),对生物多样性、经济和环境造成了严重的危害(Wanetal.,2017)。研究表明,黄顶菊能与AM真菌形成良好的共生关系,且AM真菌在黄顶菊的生长建群和入侵过程中起着重要的作用(季彦华等,2013,2015; Waggetal.,2011)。黄顶菊入侵改变了AM真菌群落的结构和多样性,即入侵地土壤AM真菌优势种发生改变,在原有的G.iranicum和G.perpusillum的基础上增加了根内球囊霉Rhizophagusintraradices(李乔,2014),且随着入侵程度的增加,G.perpusillum的孢子密度也随之增大(Zhangetal.,2017)。此外,黄顶菊根际土壤AM真菌的侵染率和生长发育与入侵地土壤理化性质密切相关(何博和贺学礼,2010),菌丝侵染率与土壤有机质呈显著正相关,与pH值呈显著负相关(季彦华等,2015)。进一步研究表明,黄顶菊根际聚集的AM真菌在竞争生长中对黄顶菊产生了偏利反馈,有助于黄顶菊的入侵(张玉曼等,2015)。吕远等(2011)发现黄顶菊与不同本地植物竞争生长时其竞争效应不同。那么,不同本地植物是否可通过改变黄顶菊根际土壤AM真菌群落结构来影响黄顶菊的竞争效应。因此,本研究采用野外同质园试验,研究黄顶菊与3种本地植物竞争生长对AM真菌群落结构和多样性的影响,获知黄顶菊根际土壤AM真菌的变化是否受本地植物种类的影响,以期从AM真菌的角度揭示黄顶菊入侵的土壤微生物学机制,同时为其生物防控提供一定的理论依据。
1 材料与方法
1.1 试验地点概况
试验地点位于河北省廊坊市广阳区九州镇中国农业科学院植物保护研究所(39°30′42″N,116°36′07″E,海拔28 m),属暖温带大陆性气候。该地土壤为棕色砂质黏壤土,土壤pH值偏弱碱性。
1.2 试验设计
试验样地自2007年开始模拟黄顶菊入侵过程,依据植株盖度设置了不同入侵程度同质园小区(李会娜等,2009):(1)黄顶菊单种处理,盖度>90%视为重度入侵区;(2)黄顶菊与本地植物的混种处理,90%≥黄顶菊盖度>60%视为中度入侵区;(3)样区内无黄顶菊,本地植物单种处理视为未入侵区。每小区长3 m,宽2 m,各小区均设置1 m的隔离带以防止边缘效应。小区主要包括本地植物单种(未入侵区)、黄顶菊与本地植物混种(中度入侵区)和黄顶菊单种(重度入侵区)3个处理,并以裸土作为对照,其中本地植物选取华北地区常见的3种伴生植物:狗尾草Setariaviridis、藜Chenopodiumalbum和黄香草木樨MelilotusofficinalisL.,狗尾草为一年生单子叶杂草,藜为一年生双子叶植物,黄香草木樨为双子叶豆科植物。各处理采用完全随机区组设计,每个处理4个重复。自2007年每个小区播种100粒种子,混种中黄顶菊与本地植物的比例为1∶1。每2周进行人工除草,保持小区植物种类不变;每年秋季拔除植物,第二年靠其种子自然繁衍生长。经过10年生长,于2016年8月份对小区植被进行调查及土样采集,调查结果见表1。

表1 小区植物株密度、生物量及盖度调查概况Table 1 AMF community parameters including plant density,biomass and coverage
1.3 土样采集
2016年8月对每个试验同质园小区土壤样品进行采样,通过形态学鉴定AM真菌种类以及测定孢子密度、种的丰度、香农—威纳指数和多样性指数等相关指标,探究黄顶菊与3种本地植物竞争对土壤AM真菌群落结构和多样性的影响。每个小区利用对角线取样法,随机选取5株生长状况良好的植物的根际土壤,单种处理采集主根根际土壤,黄顶菊和本地植物混种小区采集黄顶菊根际土壤。将所有采集的土壤过20目筛(筛网孔径830 μm),保存于-20 ℃冰箱。
1.4 AM真菌多样性调查及指标计算
采用湿筛倾注—蔗糖离心法(Phillips & Hayman,1970)分离孢子,在体视显微镜(OLYMPUS SZX16)下观察孢子形态特征,用微吸管将孢子挑取至载玻片上,加浮载剂制片,用Meelzer′s试剂进行染色并观察。参考分类标准(http:∥fungi.invam.wvu.edu/)进行孢子形态鉴定,同时参考Morton & Benny(1990)、刘润进和陈应龙(2007)分类要求,计算出AM真菌的各个多样性指标:
种数(species number,SN):某生境中AM真菌含有的物种总数;
种的丰度(species richness,SR):某一种AM真菌出现的总的次数与土样数的比值;
分离频度(frequency,F):某种真菌孢子总次数/土样数;
相对多度(relative abundanc,RA):种或属的数目/群落物种数,RA=n(某属或种的孢子数量)与N(样地分离孢子总数)的比值,以%来表示;
香农—威纳指数(Shannon-Weiner,H):H=-∑ki=1Pi×lnPi,k指AM真菌种类数目,Pi指某种孢子数/样地孢子总数;
均匀度指数(evenness index):不同种类的AM真菌在群落中的分布程度:J=H/lnS,H是指香农—威纳指数,S是指AM真菌的种类数;
重要值(important value,I)=(F+RA)/2。
本文将AM真菌的重要值划分等级(贾月月,2016):当I>60%为优势种,60%≥I>40%为亚优势种,40%≥I>20%为伴生种,I≤20%为偶见种。
1.5 数据分析
利用 Microsoft Excel 2003对数据进行初步整理,用SPASS 19.0(IBM)统计分析软件对不同处理中AM真菌孢子密度、种的丰度、香农—威纳指数和均匀度指数多样性指数进行单因素(One-way ANOVA)方差分析。
2 结果与分析
2.1 黄顶菊和狗尾草竞争对AM真菌群落结构和多样性的影响
由表2可知,黄顶菊和狗尾草3个不同处理中AM真菌种类发生变化,狗尾草单种处理分离鉴定出5属14种AM真菌,黄顶菊和狗尾草混种处理共分离鉴定出5属16种AM真菌,黄顶菊单种处理共分离鉴定出5属14种AM真菌,其中混种处理中AM真菌种类最多。
3个处理的共同优势种为网状球囊霉G.reticulatum,共同亚优势种为双网无梗囊霉Acaulosporabireticulata和摩西球囊霉F.mooseae。裸土中分离的AM真菌的优势种为网状球囊霉、根内球囊霉Rhizophagusintraradices和缩球囊霉Septoglomusconstrictum,亚优势种有双网无梗囊霉、摩西球囊霉、黑球囊霉G.malanosporum和粘质球囊霉S.viscosum。
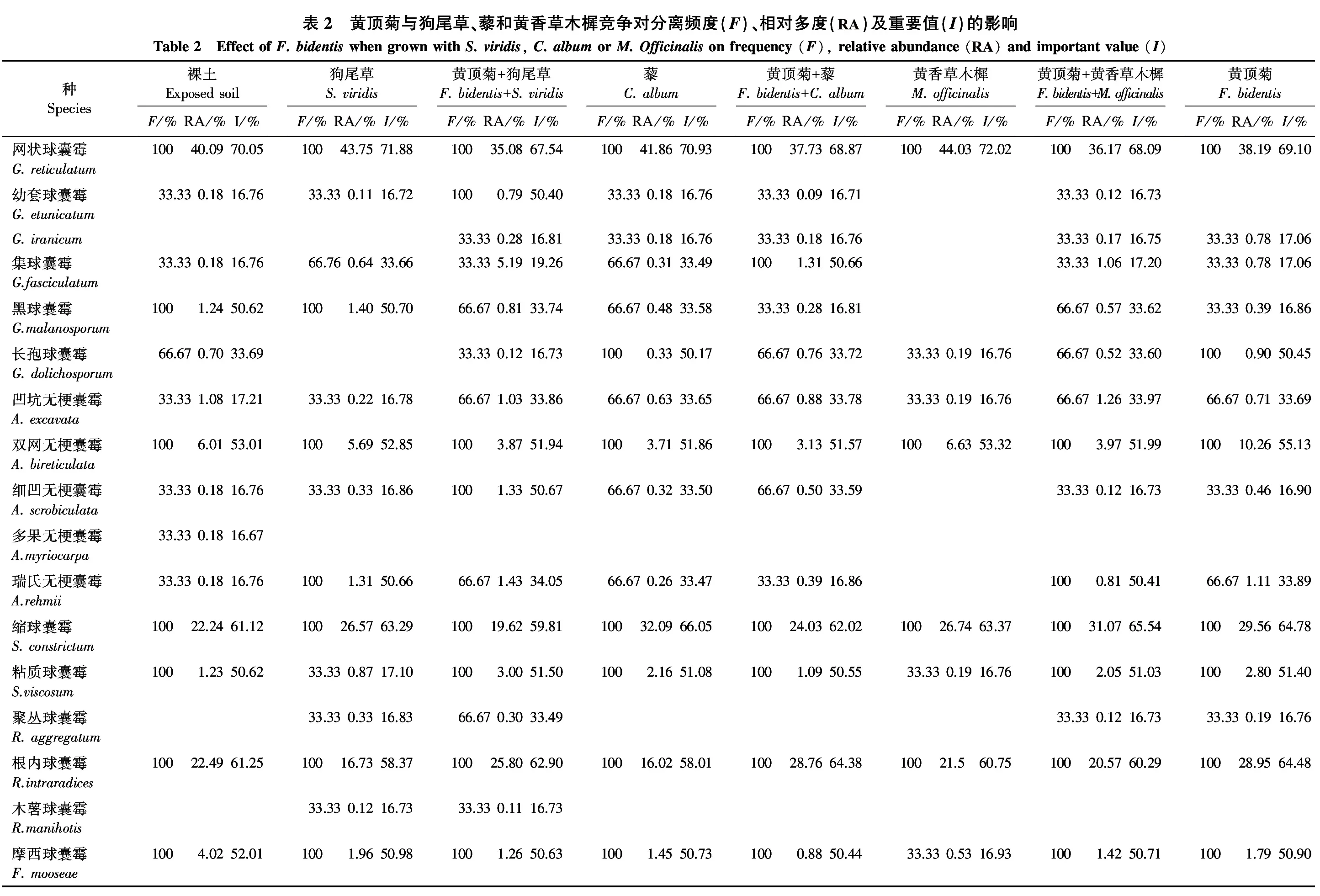
除3个处理的共同优势种网状球囊霉、共同的亚优势种双网无梗囊霉和摩西球囊霉之外,狗尾草单种处理分离得到的优势种还有缩球囊霉,亚优势种还有根内球囊霉、黑球囊霉和瑞氏无梗囊霉Acaulosporarehmii;黄顶菊和狗尾草混种处理的优势种还有根内球囊霉,亚优势种还有缩球囊霉、粘质球囊霉、细凹无梗囊霉A.scrobiculata和幼套球囊霉G.etunicatum;黄顶菊单种处理的优势种还有缩球囊霉和根内球囊霉,亚优势种还有粘质球囊霉和长孢球囊霉G.dolichosporum(图1)。与狗尾草单种处理相比,黄顶菊与之混种处理和黄顶菊单种处理中均增加了G.iranicum和长孢球囊霉2种AM真菌;此外与狗尾草单种处理相比,混种处理中根内球囊霉、粘质球囊霉、细凹无梗囊霉和幼套球囊霉的重要值增加,黄顶菊单种处理中根内球囊霉、粘质球囊霉的重要值增加。与黄顶菊和狗尾草混种处理相比,黄顶菊单种处理中减少了幼套球囊霉和木薯球囊霉Rhizophagusmanihotis2种AM真菌,根内球囊霉和长孢球囊霉的重要值增加。随着黄顶菊入侵程度的增加,黄顶菊根际土壤主要聚集了根内球囊霉和长孢球囊霉。
由表3可知,狗尾草单种处理、黄顶菊和狗尾草混种处理、黄顶菊单种3个处理中AM真菌多样性各项指标均无明显差异,说明黄顶菊入侵对AM真菌多样性没有影响。

图1 AM真菌形态学特征Fig.1 Morphological characteristics of AMFA:网状球囊霉;B:幼套球囊霉;C:黑球囊霉;D:长孢球囊霉;E:凹坑无梗囊霉;F:双网无梗囊霉;G:细凹无梗囊霉;H:缩球囊霉;I:粘质球囊霉;J:根内球囊霉;K:摩西球囊霉。A:G.reticulatum; B:G.etunicatum; C:G.malanosporum; D:G.dolichosporum; E:A.excavata; F:A.bireticulata; G:A.scrobiculata; H:S.constrictum; I:S.viscosum; J:R.intraradices; K:F.mooseae.
2.2 黄顶菊和藜竞争对AM真菌群落结构和多样性的影响
由表2可知,黄顶菊和藜3个不同处理中AM真菌的种类未发生变化,藜单种处理、黄顶菊和藜混种处理、黄顶菊单种处理中均分离鉴定出AM真菌5属14种。
3个处理的共同优势种为网状球囊霉和缩球囊霉,共同亚优势种为双网无梗囊霉、粘质球囊霉和摩西球囊霉。除3个处理的共同优势种和共同的亚优势种之外,藜单种处理的亚优势种还有根内球囊霉和长孢球囊霉;黄顶菊和藜混种处理的优势种还有根内球囊霉,亚优势种还有集球囊霉G.fasciculatum;黄顶菊单种处理的优势种还有根内球囊霉,亚优势种还有长孢球囊霉。与藜单种处理相比,黄顶菊与之混种处理中根内球囊霉和集球囊霉的重要值增加,黑球囊霉的重要值降低。与黄顶菊和藜混种处理相比,黄顶菊单种处理增加了根内球囊霉,减少了幼套球囊霉,长孢球囊霉和瑞士无梗囊霉的重要值增加,集球囊霉和细凹无梗囊霉的重要值降低。与藜单种处理相比,黄顶菊单种处理中增加了聚丛球囊霉Rhizophagusaggregatum,减少了幼套球囊霉,根内球囊霉的重要值增加,集球囊霉、黑球囊霉和细凹无梗囊霉的重要值降低。随着黄顶菊入侵程度的增加,黄顶菊根际土壤主要聚集了根内球囊霉。
由表4可知,藜单种处理时,其孢子密度显著高于与黄顶菊混种处理和黄顶菊单种处理;黄顶菊单种处理时,其香农—威纳指数显著高于藜单种处理,其均匀度指数显著高于与藜混种处理和藜单种处理。总之,与藜单种相比,黄顶菊入侵地AM真菌的孢子密度显著降低,而AM真菌的香农—威纳指数和均匀度变大。
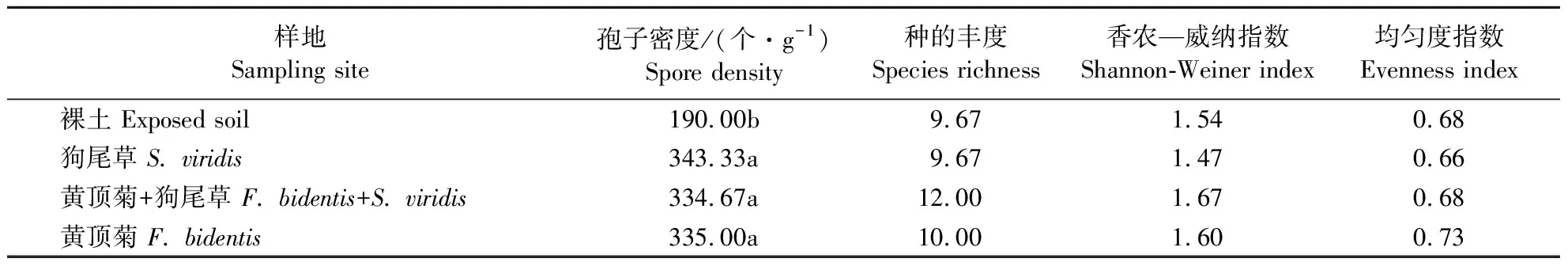
表3 黄顶菊和狗尾草竞争对AM真菌多样性的影响Table 3 Effect of F.bidentis and S.viridis community on AMF diversity index
同列数据后不同小写字母者表示差异显著(P<0.05)。
The data in the same column with the different letters indicate signficant differences (P<0.05).

表4 黄顶菊和藜竞争对AM真菌多样性的影响Table 4 Effect of F.bidentis and C.album community on AMF diversity index
同列数据后不同小写字母者表示差异显著(P<0.05)。
The data in the same column with the different letters indicate signficant differences (P<0.05).
2.3 黄顶菊和黄香草木樨竞争对AM真菌群落结构和多样性的影响
由表2可知,黄顶菊和黄香草木樨3个不同处理中AM真菌种类发生变化,黄香草木樨单种处理分离鉴定出5属8种AM真菌,黄顶菊和黄香草木樨混种处理分离鉴定出5属15种AM真菌,黄顶菊单种处理共分离鉴定出5属14种AM真菌,其中混种处理中AM真菌种类最多。
3个处理的优势种相同,即共同优势种为网状球囊霉、缩球囊霉和根内球囊霉,共同的亚优势种为双网无梗囊霉。除3个处理的共同优势种和共同的亚优势种之外,黄顶菊和黄香草木樨混种处理的亚优势种还有粘质球囊霉、摩西球囊霉和瑞氏无梗囊霉,黄顶菊单种处理的亚优势种还有粘质球囊霉、摩西球囊霉和长孢球囊霉(图1)。与黄香草木樨单种处理相比,黄顶菊与之混种处理和黄顶菊单种处理中均增加了G.iranicum、集球囊霉、黑球囊霉、细凹无梗囊霉、瑞氏无梗囊霉和聚丛球囊霉6种AM真菌,同时,长孢球囊霉、凹坑无梗囊霉A.excavata、粘质球囊霉、摩西球囊霉和根内球囊霉的重要值增加。与黄顶菊和黄香草木樨混种处理相比,黄顶菊单种处理中减少了幼套球囊霉,长孢球囊霉和根内球囊霉的重要值增加,黑球囊霉和瑞士无梗囊霉的重要值降低。随着黄顶菊入侵程度的增加,黄顶菊根际土壤主要聚集了根内球囊霉、粘质球囊霉、长孢球囊霉和摩西球囊霉。
由表5可知,黄顶菊单种处理时,其孢子密度显著高于与黄香草木樨混种处理和黄香草木樨单种处理中的孢子密度,其种的丰度和香农—威纳指数显著高于黄香草木樨单种处理;黄香草木樨单种处理时,其均匀度指数显著高于与黄顶菊混种处理。与黄香草木樨单种处理相比,黄顶菊入侵地AM真菌的孢子密度、种的丰度和香农—威纳指数显著增大,而AM真菌的均匀度指数降低。
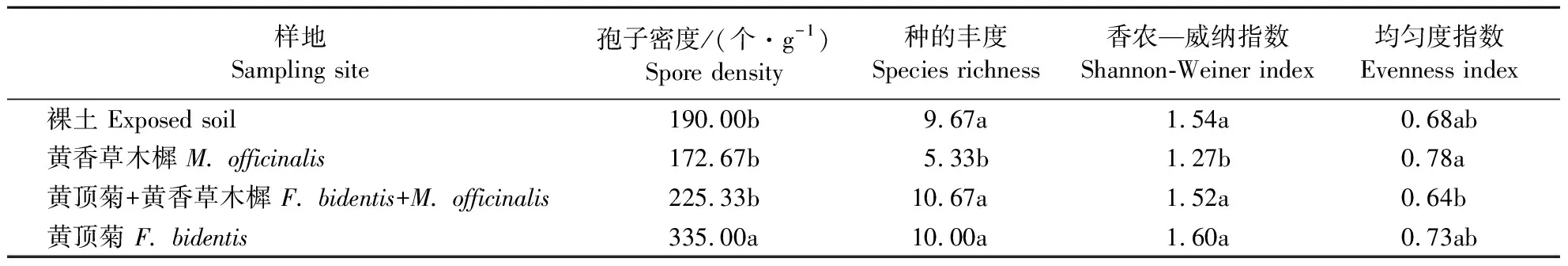
表5 黄顶菊和黄香草木樨竞争对AM真菌多样性的影响Table 5 Effect of F.bidentis and M.officinalis community on AMF diversity index
同列数据后不同小写字母者表示差异显著(P<0.05)。
The data in the same column with the different letters indicate signficant differences (P<0.05).
3 讨论
当外来植物入侵到新的生长环境中,入侵植物通过改变植物群落组成和土壤理化性质、释放化感物质等途径改变土壤微生物群落(Macketal.,2000),并改变AM真菌对原有寄主的偏好性,建立对自身有利的共生关系(Vogelsang & Bever,2009),从而快速扩张形成优势种群,影响植物多样性和生态环境。如飞机草ChromolaeneodorataL.和紫茎泽兰的入侵使AM真菌的丰富度指数显著提高(姜娜,2013);紫茎泽兰的入侵降低了隐类球囊霉Paraglomusoccultum、聚丛球囊霉和沙荒球囊霉G.deserticola的孢子密度,但显著增加了Funneliformisgeosporus的数量(Xinetal.,2013);加拿大一枝黄花入侵降低了缩球囊霉的数量,却增加了摩西球囊霉数量(Jinetal.,2004)。本试验中黄顶菊分别和狗尾草、藜、黄香草木樨3种本地植物混种处理时,黄顶菊根际土壤主要聚集的AM真菌种类不同,但均聚集了根内球囊霉。此外,在黄顶菊单种处理也聚集了根内球囊霉。已有研究表明,黄顶菊通过释放化感物质改变入侵地土壤微生物群落结构(张玉曼,2015)。黄顶菊可通过不同途径向土壤分泌黄酮和噻吩类等化感物质(彭军等,2011; 吴亚丽,2012),改变土壤养分和酶活性,从而有利于AM真菌的生长发育(何博和贺学礼,2010)。根内球囊霉能与黄顶菊形成较好的共生关系,且有助于黄顶菊竞争生长(Zhangetal.,2017),因而,可推测根内球囊霉的生长发育与黄顶菊释放的化感物质有关,相关研究有待进一步证实。
此外,入侵程度和本地植物种类是影响AM真菌群落结构和多样性的重要因素(De Vriesetal.,2012; Ylvaetal.,2013)。首先,入侵程度不同,AM真菌的多样性也不同。Marleretal.(1999)的研究表明,矢车菊Centaureamaculosa入侵到新的生长环境中与本地植物混种时,AM真菌种类增加,矢车菊快速扩张形成优势种群后AM真菌种类减少;Chenetal.(2015)研究发现,随入侵程度的增加,AM真菌的多样性和丰富度增加;Mummey & Rillig (2006)的研究表明,随矢车菊入侵程度的增加,AM真菌的群落结构发生改变,丰富度显著降低。本试验模拟黄顶菊入侵过程,人为设置了黄顶菊不同入侵程度同质园小区。黄顶菊单种处理分别与黄顶菊和狗尾草混种处理、黄顶菊和藜混种处理相比,其优势种和亚优势种的种类与数目均发生变化;黄顶菊单种处理分别与黄顶菊和狗尾草混种处理、黄顶菊和黄香草木樨混种处理相比,混种处理中AM真菌种类最多,而黄顶菊单种处理中AM真菌种类减少,且黄顶菊形成单优群落后种的丰度有所降低。该研究进一步说明AM真菌群落改变与地上植物群落结构有关(闫静等,2016),入侵程度不同AM真菌群落结构和多样性也不同。其次,已有研究表明,不同植物对不同AM真菌的菌根依赖性不同,即植物种类是影响AM真菌菌根依赖性的主要因素(初亚男等,2017;杨高文等,2015;Bardgettetal.,2015;Cantareletal.,2015;Lópezetal.,2017)。狗尾草、黄香草木樨和藜分别属于禾本科、豆科和藜科植物,3种植物根系具有不同的性状:如狗尾草根为须根系;黄香草木樨主根发达,呈分枝状胡萝卜形;藜主根发达,侧根不发达。AM真菌对3种植物的菌根依赖性和侵染率不同:AM真菌对狗尾草的侵染率为20%~25%(张玉曼,2015; 张玉曼等,2015)。黄香草木樨对摩西球囊霉有较好的依赖性,耕作土壤中摩西球囊霉对草木樨的侵染率为66.0%(肖厚军等,2013),且AM真菌能与其形成AM真菌—豆科植物—根瘤菌三者共生体系(何树斌等,2017);藜是一种非菌根植物,AM真菌侵染率很低(初亚男等,2017)。已有研究表明,不同宿主植物甚至同种宿主植物不同基因型的品种与AM真菌的共生状况不同,宿主植物的菌根长度及根中可溶性糖质量分数对AM真菌的生长发育有显著影响(陈宁等,2006);寄主类型和根系特征影响AM真菌的发生和分布,即寄主植物不同,AM真菌的侵染率、产孢率和种属构成也不同(王红新等,2006),说明植物与AM真菌之间的共生关系具有一定的选择性(Monzon & Azcon,1996)。所以,当黄顶菊分别与狗尾草、黄香草木樨和藜3种本地植物种植处理时,3种不同的本地植物导致黄顶菊根际土壤聚集不同的AM真菌,不同处理AM真菌多样性指数也不同。
综上可知,黄顶菊入侵改变了根际土壤AM真菌的群落结构和多样性,AM真菌的改变既与本地植物种类有关,也与入侵程度有关。
猜你喜欢
昆明医科大学学报(2021年12期)2021-12-30
现代仪器与医疗(2021年2期)2021-07-21
作文周刊·小学一年级版(2020年40期)2020-10-19
江苏农业科学(2017年24期)2018-01-12
环境科技(2017年1期)2017-09-05
中华骨与关节外科杂志(2017年1期)2017-05-17
农业与技术(2017年6期)2017-04-21
小学生作文选刊·低年级版(2016年6期)2016-06-24
微创泌尿外科杂志(2014年4期)2014-02-28
中国火炬(2012年9期)2012-07-25
