
垂体特异性转录因子与家畜经济性状关系的研究进展
2018-09-08 05:27:26李佳蓉姜怀志
家畜生态学报 2018年8期
李佳蓉,贾 超,姜怀志
(吉林农业大学,吉林 长春 130118)
POU结构域因子家族是一类转录调节因子,其家族特点是DNA结合结构域(即POU结构域)具有高度保守性。而“POU”即来源于垂体特异性转录因子Pit-1[1]、八聚体结合蛋白Oct-1和Oct-2及caenorhabeditⅠ线虫神经Unc-86[2]这4种转录因子英文名字的首字母。垂体特异性转录因子(pituitary specifific transcription factorl,POU1F1;也称为PIT-1或GHF-1),是POU同源异型域转录因子家族的成员之一。POU1F1最初被认为是一种生长激素因子,后经研究表明, 它是一种组织特异性转录因子。Ohta等在1992年首先将人类的POU1F1基因定位在第3号染色体的3p11区域[3]。该基因全长约14 kb,其mRNA全长1 262 bp,编码291个氨基酸[4]。Woollard等[5]于2000年利用原位杂交技术,将牛、绵羊和山羊的POU1F1基因定位于1q21-22染色体上,主要功能是调节细胞的增殖与分化、调控动物生长发育。
1 POU1F1的结构与生物学特性
垂体特异性转录因子POU1F1是一种在脑垂体中对多个基因的转录调控中具有关键作用的转录因子,是在下丘脑核群中产生的蛋白因子。作为POU结构域家族基因的成员,POU1F1(也称为PIT-1或GHF-1)主要表达于动物的脑垂体前叶细胞和增殖、生长的三种腺瘤细胞类型[6],并影响其表达的正常分化。它的主要功能一是调节细胞分化、调控动物生长发育,能够识别特异的基因顺序并与之结合,从而引起细胞内基因的转录;二是调节三种垂体前叶细胞(甲状腺细胞,生长激素和催乳素细胞)以及生长激素(GH)、催乳素(PRL)、促甲状腺激素(TSH)基因和POU1F1基因的正常分化、增殖和生长[7],并促使以上基因的转录和表达,从而影响动物的生长、发育和繁殖[7-9]。
国内外大量研究结果表明,小鼠POU1F1的突变与Snell矮小(dw)和Jackson矮小(dw-J)相关联,人的POU1F1突变也被发现为与之相关[6,10-11]。牛、绵羊和山羊的POU1F1基因已定位于lq21~22区[5]。研究表明,在此区域内,猪、牛POU1F1基因的突变与生产性状的显著关系和QTL定位都揭示了POU1F1基因能影响动物的生产性能[12-14]。
2 POU1F1在绵、山羊上的相关研究
Bastos等[15]首先揭示羊POU1F1基因的多态性,初步阐述山羊POU1F1基因突变及其与生产性状的关系。Lana等[9]首次报道了山羊POU1F1基因座中一个沉默等位基因的鉴定,并对801个样品中的AluI PCR-RFLP进行了检测,初步阐述山羊POU1F1基因突变及其与生产性状的关系。此外,还对山羊POU1F1突变与生产性状之间的关系进行了评估。不同类型与乳产量(P<0.05)和出生体重(P<0.05)的显著关联意味着POU1F1基因对产奶性能和早期体重有积极的影响。山羊POU1F1基因对毛发性状没有显著影响。因此,Bastos等首次提出基因型TC和TT分别被认为是优质奶产量和出生体重的分子标记。
Ozmen等[16]检测了三种土耳其绵羊品种(Sakiz,卡拉曼白羊,Awassi绵羊)的POU1F1基因外显子6和3'UTR区域内的AluⅠ和DdeⅠ多态性及其与羊奶生产性状的关系。经研究后发现限制性内切核酸酶AluI具有双重多态性:TT(296 bp和173 bp)和CC(235 bp,173 bp和61 bp),而没有检测到DdeI切割位点的多态性。在TT和CC基因型个体中,乳产量(P<0.001)、脂肪(P<0.001)和乳糖(P<0.05)含量差异显著,蛋白质含量(P>0.05)差异不显著。在Sakiz绵羊品种中基因型TT个体具有优异的产奶性能。测序结果表明,POU1F1基因外显子6(g.185T>C)和3'UTR(g.220G>A,g.229C>T,g.248C>T,g.250A>T,g.255T>C, g.258C>T)共存在七个变异点。
羊POU1F1基因位于染色体1上,含有5个内含子和6个外显子。在不同品种的哺乳动物中不同外显子的突变与不同的生产性状有关。Mura等[17]研究检测POU1F1基因核苷酸序列多态性及其与Sarda羊奶生产性状的关系。运用PCR-SSCP方法对6个外显子进行了测序,分析了可能的SNP。分析变体与产奶量的关系和其组成后发现了两种新型SNP:在第四个内含子片段中的5'UTR有121 C>T和在第六外显子片段中的3'UTR有249 G>A。统计分析发现结果未显示羊奶生产性状之间的关联和多态性。绵羊的POU1F1基因具有高水平的保守性,其与牛、人和鼠的同源性分别为98.2%,91.2%和86.2%。Bastos等[15]对葡萄牙本地羊品种"Churra da Terra Quente"的POU1F1进行测序,发现测序的5 787 bp包括六个外显子和两个完整的内含子,并分析检测这六个外显子多态性。在外显子2的58位密码子中发现一个多态性,一个等位基因变化导致了半胱氨酸变为酪氨酸(2%等位基因频率)。在外显子3中,检测到两个多态性:在89密码子处的G→A转变为甘氨酸→天冬酰胺(0.5%等位基因频率),在105密码子处的G→A转变为丙氨酸→苏氨酸(1.5%等位基因频率)。这些多态性可能会改变POU1F1蛋白的结构并修饰基因的表达。在内含子4区域检测到了一个A→G转变(3%等位基因频率)。而外显子1,4和6此次没有检测出多态性。
王玺年等[18]采用PCR-SSCP和DNA测序技术,对河西绒山羊、陇东绒山羊、辽宁绒山羊和内蒙古绒山羊4个群体共509个个体的POU1F1基因第3外显子和第6外显子进行SNP检测和遗传多样性、生产性状遗传效应分析,以期探寻山羊分子育种的有效遗传标记。结果显示:(1)在POU1F1基因第3外显子中检测到1个SNP位点 (c.313A>G)并形成AA和AB 2种基因型,第6外显子中检测到2个SNPs位点( c.682G>T、c.837C>T)并形成AA, BB, AB, AC和BC 5种基因型。(2)遗传多态性分析表明,4个山羊群体在第3外显子区域处于低度多态(PIC<0.25),在第6外显子区域处于中度多态(0.25
罗卫星等[19]研究采用PCR测序技术对黔东南小香羊和贵州黑山羊的POU1F1基因外显子6进行SNP筛查,并分析其与屠宰性状的关联性。结果表明:在2个山羊群体POU1F1基因外显子6均检测到2个SNPs(T58G和T172C);在黔东南小香羊群体中,POU1F1基因外显子6的 T58G位点AA和aa型个体的屠宰率显著高于Aa型(P<0.05),T172C位点Bb型个体的屠宰率显著高于BB和bb型(P<0.05);贵州黑山羊群体中,T58G位点Aa型个体的胴体重、屠宰率、眼肌面积厚和腰部肌肉厚显著低于AA和aa型(P<0.05),瘦肉率显著低于AA型(P<0.05),T172C位点BB型个体的胴体重、屠宰率、眼肌面积、腰部肌肉厚、净肉重、瘦肉率和骨重显著低于Bb和bb型(P<0.05);其他各性状在各位点各基因型之间差异未达到显著性差异(P>0.05),由此推测POU1F1基因外显子6的多态可能对山羊的屠宰性状有显著的遗传效应。
Li等[20]研究描述了15种中国山羊POU1F1-DdeI基因座的多态性并分析了其中等位基因的分布情况。 PCR-RFLP分析显示D1D1和D1等位基因的优势基因型,平均频率为0.550和0.790,不分山羊的效用类型。D1D2基因型最频繁,平均频率为0.371。基于卡方检验发现,该基因座位和基因型等位基因频率分布差异显著(P<0.001),表明品种因子显著影响POU1F1基因的分子遗传特征。遗传多样性分析显示,中国土著种群在山羊POU1F1-DdeI基因座上具有广泛的遗传多样性。然而,ANOVA分析显示,肉类、乳制品和羊绒类产物类型中基因纯合性、基因杂合度、有效等位基因数量和多态性信息含量无显著性差异(P>0.05)。这表明山羊效用类型对遗传多样性没有显著影响。
在此研究中,Jalilsarghale等[21]研究了POU1F1基因第三外显子的单核苷酸多态性(SNPs)及其与两种伊朗羊品种Zel和Lori-Bakhtiari的生长特征和血液代谢物的关系。收集90只Lori-Bakhtiari和90只Zel羊的血液样本,提取DNA,扩增POU1F1基因的295 bp片段,采用限制性片段长度多态性(RFLP)技术进行基因分型。在Lori-Bakhtiari和Zel绵羊品种中鉴定出SNP,其代表在内切核酸酶AciI的限制性位点处的非同义单碱基突变。结果显示两个研究品种之间等位基因的差异频率,其中A等位基因在Lori-Bakhtiari品种中更为频繁,而G等位基因在Zel品种中更为频繁。当检测到POU1F1基因型时,AA基因型的动物具有比GG基因型的动物更高的断奶重量(P<0.05),然而基因型与出生体重之间,生物特征(体长,体高,心脏周长,大腿围和腹围)和血液代谢物(甘油三酯和胆固醇)之间(P>0.05)没有显著的关联。这些发现意味着POU1F1多态性可能影响断奶重量,因此可用作该生产性状的分子标记。
赵宪林等[22]为探讨POU1F1基因的多态性与藏羊生长性状之间的关联性,寻找与藏羊生长性状相关的分子标记。利用DNA测序方法检测藏羊POU1F1基因的多态位点并对其进行基因分型,分析单一多态位点不同基因型及双倍型与藏羊生长性状的关联性。结果显示在POU1F1基因第4内含子上检测出1个多态位点g.14205G>A,在第5内含子上检测出2个多态位点g.15223G>A和g.15286A>G。关联性分析表明,g.14205G>A位点上AA基因型的体质量和体长分别极显著(P<0.01)和显著(P<0.05)高于GG和AG基因型;g.15223G>A位点上GG基因型的体质量显著高于GA基因型(P<0.05),极显著高于AA基因型(P<0.01);g.15286A>G位点AA基因型体质量极显著高于GG基因型(P<0.01),体高显著高于GG基因型(P<0.05)。单倍型Hap7(-GGA-)的发生频率最高,达到42.40%,其次为单倍型Hap5(-GAA-)和单倍型Hap3(-AGA-),发生频率分别为21.50%和14.70%。此外,POU1F1的r2值均小于0.33,说明这3个多态位点之间不存在强的连锁性。通过合并基因型发现,双倍型H7H7(GGA-GGA)的各个生长性状表现最优,其个体的体质量、体长和体高大于其他双倍型。说明POU1F1基因的3个SNP位点均能够显著影响其生长发育,优势基因型分别为AA、GG和AA,可用于藏羊生长性状的选育。
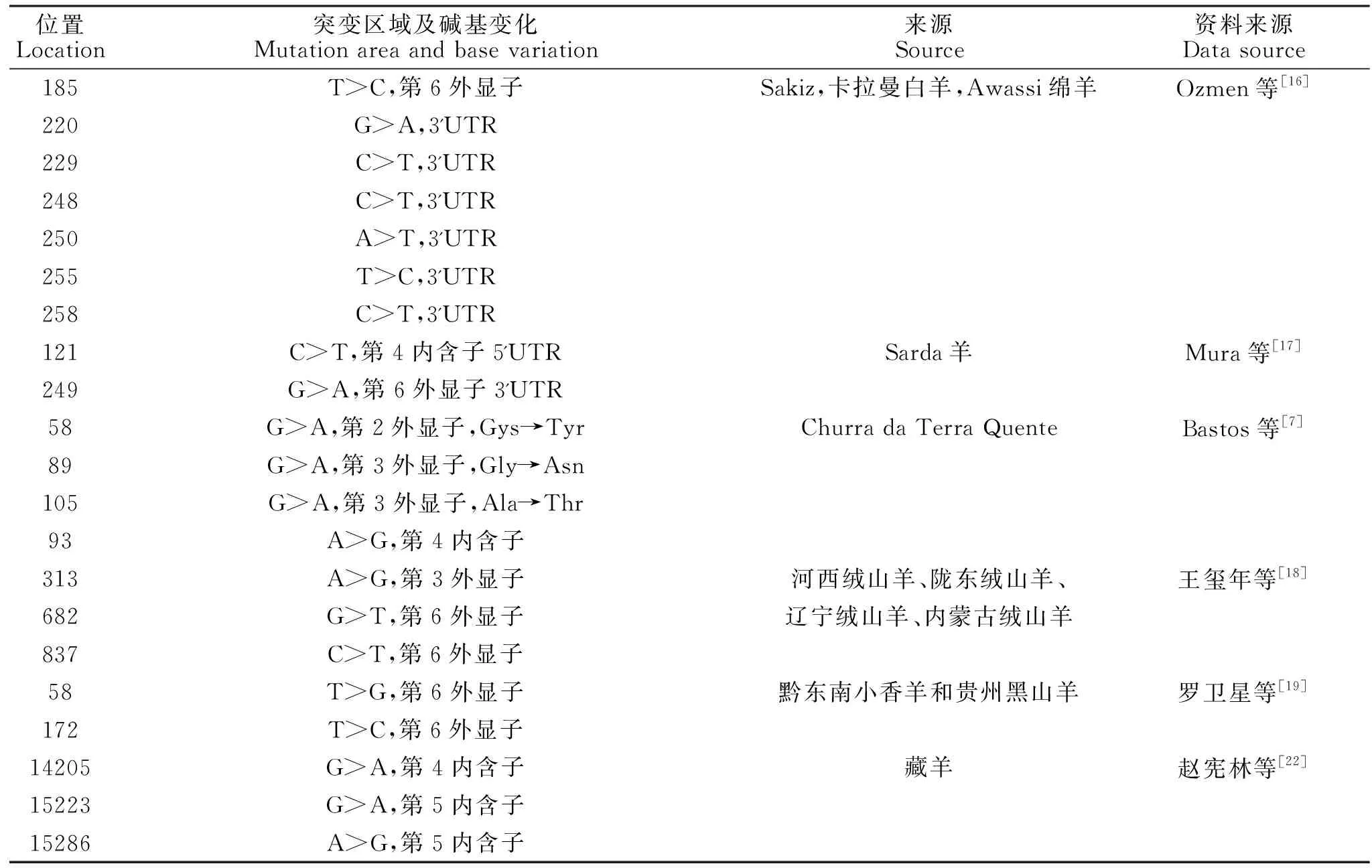
表1 羊POU1F1基因的突变Table 1 Mutations of POU1F1 gene in sheep
3 POU1F1在牛上的相关研究
在牛中,位于POU1F1基因第6外显子的Hinf1的多态性对奶量有显著影响。结果显示A等位基因与较高的产奶量相关,并且可能是改善牛奶量的遗传标记。为了更好地了解这种多态性对牛的乳腺发育和功能的影响,Carsai等[23]研究了罗马尼亚黑牛、罗马尼亚白牛品种的多态性,前一种是与罗马尼亚灰色草原品种相比具有高产奶性能的牛品种,后一种是产奶量非常低的原始品种。在两个品种中,B的等位基因频率比A的等位基因频率高。
Xue等[24]应用PCR-RFLP分析POU1F1基因遗传变异对100只南阳牛生长性状的影响。结果表明,用HinfⅠ消化的451 bp PCR产物在牛群中表现出多态性,处于Hardy-Weinberg平衡状态。南阳牛群的A/B等位基因频率分别为0.465/0.535。分析POU1F1基因变异与群体生长性状的关系后发现,具有基因型BB的个体内以下参数比具有基因型AB的个体更优秀:出生体重、断奶前平均增重量、12月龄时体高、体重、体长和6月龄和12月龄的胸围(P<0.05)。BB个体12月龄体重高于AA组(P<0.05)。其他年龄组的体重和体型也呈现等位基因B>等位基因A的趋势。因此,基因型BB可能是显性基因型,等位基因B可能是显性等位基因。这些结果意味着POU1F1基因的等位基因B可能对生长特性具有积极的影响。
Myogenic因子5(Myf5)和POU1F1基因在哺乳动物的生长和发育中起重要作用。利用牛Myf5和POU1F1的特性来检测遗传变异位点,并将它们与367头牛(Hanwoo(325)和Angus(37))的经济性状进行关联分析。Seong等[25]通过基因组DNA的序列分析,鉴定出Myf5的内含子2(A1948G SNP)和POU1F1的外显子6(A15635G SNP)中的两个单核苷酸多态性(SNP)。统计学分析显示,Myf5多态性与6个月龄的背脂厚度和活体重显著相关,POU1F1多态性显著影响24个月龄胴体重和体重、背脂肪厚度。Myf5和POU1F1之间的相互作用在胴体重、背最长肌面积、背脂肪厚度和大理石花纹分数方面均显著。表明Myf5和POU1F1作为生长和胴体性状的候选基因,它们之间的相互作用显著影响牛的生长和胴体性状。
陈然等[26]运用PCR-RFLP技术研究了新疆褐牛266头个体POU1F1基因的多态性,在新疆褐牛群体中发现A、B两种等位基因,并发现了AA、AB、BB 3种基因型。相应得到等位基因A/B频率、基因型的频率、纯合度(H0)、杂合度(He)、有效等位基因数(Ne)、多态信息含量(PIC)的统计数据。结果表明:新疆褐牛POU1F1基因座的451bpPCR产物经限制性酶HinfI消化后表现多态,其等位基因A/B频率为0.3083/0.6917;AA、AB和BB3种基因型的频率分别为:0.0677/0.4812/0.4511,在该基因座处于Hardy-Weinberg平衡(P>0.05);它们在该基因座的纯合度/杂合度/有效等位基因数/多态信息含量分别为:0.5735/0.4265/1.74/0.3356。这表明了新疆褐牛在人工选育、迁徙和遗传漂变等因素作用下POU1F1基因座处于动态平衡状态;并得出A等位基因在群体中不具有优势,A等位基因的劣势效应可以作为标记辅助选择(MAS)的参考,同时也给育种工作者建议应加大改良力度,对早期选种有一定指导意义。
西藏牦牛POU1F1基因存在一定的遗传多态性,宋娜娜等[27]利用DNA池和测序技术,结合PCR-RFLP方法,筛查了POU1F1基因第2外显子和第5内含子的SNP位点,分析了西藏申扎(SZ)、类乌奇(LWQ )、斯布(SB)和帕里(PL)4个牦牛类群(品种)共182个个体的POU1F1基因的多态性及其与体重、体高、体长等生长性状指标之间的相关性。结果表明:(1)POU1F1基因第2外显子高度保守,未发现任何突变,第5内含子中发现2个突变位点,分别为T727C (BgllⅠ酶切)位点和T284C(StuⅠ酶切)位点。(2)申扎、类乌奇、斯布和帕里4个群体中StuⅠ位点为中度多态性,其基因T/C频率分别为:0.61/0.39、0.54/0.46、0.54/0.46和0.62/0.38,BgllⅠ位点为低度多态性;Hardy-Weinberg检验表明StuⅠ位点和BgllⅠ位点均处于平衡状态(P>0.05)。(3)StuⅠ位点在4个群体中表现CC、CT和TT 3种基因型,Ⅱ位点在4个群体中只表现CT和TT2种基因型。(4)帕里牦牛StuⅠ位点CC和CT基因型在生长性状上存在显著差异(P<0.05),其中CC基因型的体高和体长均高于CT基因型,而其他几个牦牛类群末发现类似现象。BgllⅠ位点基因型与生长指标的相关性不显著。西藏牦牛POU1F1基因内含子5内T284C基因座表现的多态性对帕里牦牛的体高和体长有较为显著的影响,可以作为改良其生长性状的一个遗传标记。
4 POU1F1在猪上的相关研究
人们分离了猪POU1F1部分cDNA和其整个3'UTR的基因组克隆,发现POU1F1的多态性与猪的几种性状特征相关。为了了解猪POU1F1基因的生物学功能及其在猪遗传学中的应用,Yu等[28]使用逆转录酶-聚合酶链反应(RT-PCR)进行猪POU1F1全长cDNA的克隆。猪POU1F1基因的cDNA及其推导的蛋白质序列分别与其他哺乳动物(人,牛,绵羊和啮齿动物)的POU1F1cDNA和蛋白质具有约90%~95%的同源性。但是与其它猪的POU1F1序列相比较只显示出约93%的同一性。Yu等[12]对POU1F1基因进行了测序并在外显子4中鉴定了一个新的多态性。几种哺乳动物POU1F1序列的进化分析表明,测序误差可能解释了其他猪的序列报告中观察到的差异。还通过RT-PCR鉴定了几种POU1F1选择性剪接形式。它们是△3POU1F1(缺少整个外显子3),△4POU1F1(缺少整个外显子4)和POU1F1β(插入外显子2前面的26个氨基酸)转录物。在啮齿动物中发现了△4POU1F1和POU1F1β转录物编码功能上的差异蛋白质。△3POU1F1转录本是POU1F1的新亚型。通过细菌中这些蛋白质的表达来分析猪△3POU1F1和POU1F1之间潜在的不同功能。在DNA迁移率变动测定中大肠杆菌表达的POU1F1和△3POU1F1蛋白与大鼠生长激素(rGH)和大鼠催乳素(rPRL)启动子DNA共同作用。结果表明,猪POU1F1可以特异性结合rGH和rPRL启动子区域,但即使是在非常高的蛋白质浓度下猪△3POU1F1也不能与之结合。在凝胶移位测定前通过混合蛋白质提取物测试△3POU1F1和POU1F1之间的蛋白质的相互作用,结果表明△3POU1F1蛋白不影响POU1F1与靶DNA的结合。这些数据证明了此研究中克隆的POU1F1cDNA的功能,并鉴定了一种新型△3POU1F1转录物,其不能编码结合rGH/rPRL靶序列的蛋白质。
中国梅山猪在初生后100 d内的发育期发育迅速,与英国/欧洲的品种相比胎盘尺寸较小,产仔数较大。Sun等[14]为了研究POU1F1在猪生长和繁殖性状中的作用,对出生后第0,15和30天的9窝仔猪采集垂体(n=60)和/或血液样品(n=80),并将所有动物进行基因分型(CC,CD,DD)用于分析研究MspⅠPOU1F1的多态性。研究结果表明出生时POU1F1基因型对GH和PRL的循环水平有显著影响。DD基因型的仔猪GH水平较低,但PRL水平高于其他基因型。POU1F1-α的mRNA从0天到第30天下降(P<0.05),与同期相比GH的mRNA以及PRL和GH血浆水平平行减少(P<0.05)。POU1F1-βmRNA水平在这一时期并没有显著变化。POU1F1-αmRNA与GH mRNA和GH在血浆浓度水平之间以及两种POU1F1 mRNA同种型之间的相关性是显著的。
张陈华等[29]采用PCR-RFLP (RsaⅠ限制性内切酶)分子标记技术,利用现代分子标记技术寻找影响猪生长性状的标记基因座,分析了杜洛克、长白、大白及长大二元杂交4个品种共381头猪POU1F1基因外显子 5至内含子5内多态性,并采用最小二乘法分析了POU1F1基因多态性对生长性状影响的遗传效应。经检测,4个品种POU1F1基因RsaⅠ位点均存在多态性。等位基因A的基因频率分别为0.8317、0.8302、0.6767、0.7523,A为优势基因。被测猪群POU1F1基因的基因频率在被测猪群中均处于 Hardy-Weinberg平衡状态(P>0.05)。最小二乘分析表明:POU1F1基因3种基因型的猪在出生重、断奶重及70日龄重性状上均呈现BB>AB>AA趋势(其中,长白猪只存在AB和AA 2种基因型),B为优势基因。POU1F1基因外显子 5至内含子5 RsaⅠ位点多态性对测试猪群生长性状有一定影响,但是否是影响生长性状的一个候选基因之一或是与影响猪生长性状的QTL连锁的基因,还需进一步研究。
许金根等[30]为寻找改良猪生长性能的分子标记,研究猪POU1F1基因多态性及其对生长性状的相关性。采用PCR-RFLP方法检测猪POU1F1基因的单核苷酸多态性(SNP),并分析基因多态性与生长性状的关联性。结果显示猪POU1F1基因的RsaI酶切位点存在多态性。其中,杜洛克猪未检测到AA型,BB基因型频率高于AB型,等位基因B频率高于等位基因A,SNP处于低度多态(PIC<0.25);淮猪新品系未检测到BB型,AA基因型频率高于AB型,等位基因A频率高于等位基因B,SNP处于低度多态(PIC<0.25);杜洛克猪×淮猪新品系未检测到BB型,AB基因型频率高于AA型,等位基因A频率高于等位基因B,SNP处于中度多态(0.25
而YU等[31]研究发现,POU1F1基因的RsaI酶切位点在枫泾猪、梅山猪和民猪仅检测到AA型,切斯特白猪、杜洛克猪和汉普夏猪检测到三种基因型,长白猪和大白猪只检测到AA和AB型 。另外,庞瑾等[32]研究检测POU1F1基因的RsaI酶切位点在大白猪、长白猪和杜洛克猪结果显示,三个猪种均存在处于中度多态的AA、AB和BB基因型,且都以基因型AA频率最高和等位基因A频率较高。BRUNSCH等[33]研究发现POU1F1基因RsaI酶切位点多态性与欧洲野猪×皮特兰猪的F2代14个生长和胴体组成性状显著相关,但与梅山猪×皮特兰猪的F2代所测性状无显著相关,这说明不同猪群POU1F1基因RsaI酶切位点对生长性状的遗传效应可能不一样。因此,这些试验结果不一致,除了还需扩大样本进行研究,可能与猪的遗传背景差异有关。
5 小结与展望
POU1F1作为在下丘脑核群中产生的一类转录调节因子,它的突变可能导致GH,PRL,TSH和POU1F1基因本身不同的表达从而影响这些基因的分化、增殖和生长,进而影响动物的生长、发育和繁殖。目前的研究发现POU1F1对哺乳动物的产奶性状、早期体重、生产性状、屠宰性状和胴体性状具有重要作用。Lana等[9]发现在山羊中POU1F1对毛发性状没有显著影响,但王玺年等[18]发现其对山羊产绒量存在一定影响,并认为其可作为山羊生产性状的候选基因座之一。另外,品种因子显著的影响POU1F1基因的分子遗传特征。综上所述,可看出POU1F1对机体生长发育具有重要意义,这可以有效的帮助我们在遗传育种、遗传变异的研究中提供研究思路和工作帮助。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29 01:57:52
系统工程学报(2021年4期)2021-12-21 06:21:08
智慧健康(2021年17期)2021-07-30 14:38:32
中国生殖健康(2020年4期)2021-01-18 02:58:10
新世纪智能(高一语文)(2020年12期)2020-06-01 08:14:20
中国生殖健康(2018年4期)2018-11-06 07:12:16
现代检验医学杂志(2016年5期)2016-08-20 03:16:54
法医学杂志(2015年4期)2016-01-06 12:36:40
中国医药导报(2015年27期)2015-02-28 22:08:01
湖北农业科学(2014年11期)2014-09-10 18:06:07
