
Contrasting arthropod communities associated with dwarf mistletoes Arceuthobium globosum and A.vaginatum and their host Pinus hartwegii
2018-09-07 03:06LeChvezSalcedonicaQueijeiroBolaosctorpezmezZenCanoSantanaBlancaMejRecamierAureaMojicaGuzm
Journal of Forestry Research 2018年5期
León F.Chávez-Salcedo·Mónica E.Queijeiro-Bolaños,4·Víctor López-Gómez·Zenón Cano-Santana· Blanca E.Mejía-Recamier ·Aurea Mojica-Guzmán
Abstract Arthropod communities and epiphytic plants associated with tree canopies have been widely studied and have revealed a great diversity of organisms;however,the community hosted by parasitic plants,such as dwarf mistletoes,remains poorly known.In the coniferous forests of North America,dwarf mistletoe infection(Arceuthobium spp.)significantly damages the health of the forest,causing large financial losses for the forest industry,but it also positively affects diversity,especially of mammals and birds.This study examined the attributes of the arthropod communities associated with two species of dwarf mistletoe[Arceuthobium globosum Hawksw.&Wiens and A.vaginatum(Humb.&Bonpl.ex Willd.)J.Presl]and their host Pinus hartwegii Lindl.In 2010,in five sites located in Zoquiapan(Central Mexico),we collected plant tissue from the three species bimonthly.Arthropods were separated from the plant tissue and identified to the finest level possible.We collected 32,059 individuals,for which 51 morphospecies were identified, belonging to 15 taxonomic orders;the most abundant orders for the three plants were Prostigmata,Thysanoptera and Homoptera.The community associated with P.hartwegii had the highest value of diversity(H′=1.47;A.globosum,H′=0.64;A.vaginatum,H′=0.68)and species richness(S=40;A.globosum,S=30; A.vaginatum,S=35);while abundance was significantly higher for the mistletoes(A.globosum,n=407 individuals/sample;A.vaginatum,n=536 individuals/sample;P.hartwegii,n=134 individuals/sample).Species richness,abundance and diversity were significantly different for the three studied plants,as well for sampling month and the interaction of these two factors(except for diversity).The results suggest that the canopy of P.hartwegii is an important element in the ecosystem,providing a mosaic of resources and conditions to the associated fauna.We also propose that mistletoes are key species within the forest canopy,as they greatly influence the establishment of diverse organisms,particularly arthropods.
Keywords Arceuthobium spp.·Pinus hartwegii·Arthropod community·Thrips·Mites·Hemiparasitic plants
Introduction
Insect–plant interaction is one of the most studied biotic relationships because it includes two major biological groups:plants,which represent the largest biomass source on Earth(Schoonhoven et al.2005);and arthropods,which comprise most of the living taxa(Borror et al.1989).Arthropods play important roles in plant community dynamics,acting as herbivores, pollinators and seed dispersers(Samways 1994),and plants offer food and shelter for these organisms(Schoonhoven et al.2005).The attributes of the arthropod community within a host plant are influenced by the plant’s features such as size,structural complexity,phenology(Lawton 1983;Langellotto and Denno 2004;López-Gómez and Cano-Santana 2010;Schowalter 2011),food availability,primary productivity,presence of prey(Polis et al.1997;Mittelbach et al.2001),environmental conditions(Schowalter 2011)and presence of key species(Power et al.1996;Letourneau and Dyer 1998).
Forest canopies are characterized by a high diversity and abundance of organisms,including a great variety of arthropods(Southwood et al.1982;Stork 1991;Basset and Arthington 1992;Tovar-Sánchez et al.2003)because of the great variety of exploitable resources and conditions(Morecroft et al.1985;Fitzjarrald and Moore 1995),including a vast number of epiphytic and parasitic plants that increase the environmental complexity and provide sites for colonization(Shaw 2004).Some studies have reported that epiphytic plants contain a large variety of microarthropods,mainly mites(Acari)and springtails(Collembola)(Yanoviak and Nadkarni 2001;Yanoviak et al.2003).Nevertheless,little attention has been paid to an important component on forest canopies and its interaction with invertebrate organisms:mistletoes.
Mistletoes have long been treated as pests on forestry lands because of their negative effects on hosts performance,such as decreased growth and fecundity(Mathiasen et al.2008),but are now known to play key role in their biological community by improving ecosystem processes such as decomposition and nutrient cycling(March and Watson 2007;Ndagurwa et al.2014)and enhancing vertebrate and invertebrates diversity(Watson 2001;Watson and Herring 2012).
Dwarf mistletoe infection(Arceuthobium spp.,Viscaceae)of coniferous trees is worrying worldwide because of their negative impacts on economic and forestry activities(Stevens and Hawksworth 1984;Hawksworth and Wiens 1996).In Mexico,dwarf mistletoe infection is considered the second leading cause of forest damage,after fires(Hawksworth 1983).The biology,physiology and systematic of dwarf mistletoes are well documented(Hawksworth and Wiens 1996;Mathiasen et al.2008);however,other important ecological aspects,such as the relationship with arthropods,have received less attention.
Mistletoes can be tightly associated with arthropods,since many species can act as pollinators(Coleoptera,Diptera,Hymenoptera,and Lepidoptera;Pen field et al.1976;Whittaker 1984;Hawksworth and Wiens 1996),and others can feed on its tissues(Hawksworth and Wiens 1996).Nevertheless,few studies have focused on the arthropod community held within mistletoes(Whittaker 1984).Burns et al.(2011)found 10 arthropod orders inhabiting the box mistletoe(Amyema miquelii)and its host(Eucalyptus spp.),which had very similar arthropod community compositions,but eucalyptus had greater densities of arthropods.The most representative orders were,in order of abundance,Hemiptera,Thysanoptera,Coleoptera,Araneae,Hymenoptera and Psocoptera.Results from other studies of arthropods communities on mistletoes,which did not include any host comparisons,are very similar(Room 1972;Whittaker 1984;Tassone and Majer 1997;Anderson and Braby 2009)and include the presence of other orders such as Collembola,Coleoptera and Lepidoptera(Anderson and Braby 2009).
However,to our knowledge,no study has been published for the community associated with dwarf mistletoes(Arceuthobium spp.),although various studies report single species associations.Several mite species(Prostigmata and Mesostigmata)have been described as tightly associated with different Arceuthobium species(Stevens and Hawksworth 1970;Kennett 1963;Pritchard and Baker 1958;Mathiasen et al.2008),and larvae of a phytophagous lepidopteran as a specific consumers(Mooney 2001;Larsen et al.1995).
The goal of this study was to compare the arthropod community attributes(species richness,diversity,abundance and composition)among three plant species:two dwarf mistletoes[Arceuthobium globosum Hawksw.&Wiens and A.vaginatum(Humb.&Bonpl.ex Willd.)J.Presl]and its host plant(Pinus hartwegii Lindl.)in the Zoquiapan region of Central Mexico.
Increasing our knowledge about the arthropod community associated with dwarf mistletoes and their pine host will contribute to a better understanding of arthropod–plant interactions and the role of dwarf mistletoes in these communities,other than the traditional concept of these organisms as harmful pests.Even more,the understanding of the pine-dwarf mistletoe system may be useful to plan managing strategies when needed by detecting phytophagous insects associated with dwarf mistletoes and using them for biological control.
Materials and methods
Study area
The study was conducted in the Zoquiapan subregion within the Iztaccíhuatl-Popocatépetl National Park in Central Mexico(19°15–29′N,98°37–45′W).The area occupies 19,400 ha and has a subhumid temperate climate with summer rains;the mean annual temperature is 9.7°C,and the annual precipitation is 941 mm(Arriaga et al.2002;SMN 2013).The vegetation types found in this area arePinus,AbiesandQuercusforests;nonetheless,the flora is mainly composed of Pinus hartwegii(Obieta and Sarukhán 1981).Other tree species,such as P.montezumae,P.ayacahuite,Cupressus lusitanica and Alnus jorullensis,occur at low frequency.Forest understory is represented by tussocky grasses such asMuhlenbergia macroura,M.quadridentataandFestuca tolucensis.The rainy season is from May to October,and the dry season is from November to April(Obieta and Sarukhán 1981).
Methods
Study system
Arthropod communities in association with two dwarf mistletoe species(A.globosumandA.vaginatum)and their host(Pinus hartwegii)were described.Arceuthobium globosumsubsp.grandicaule(Viscaceae)is a highly branched parasitic shrub,20–70 cm tall,with yellow or greenish stems;its staminate flowers are 5 mm wide.Anthesis was recorded between January and May,with a peak from March to April.Fruits ripen from July to October,and seeds are dispersed from July to November,with a peak in August and September.This plant parasitizes 12 species of Mexican pines;it is distributed from central Mexico to Guatemala and Honduras,within an altitudinal range of 2450–4000 m a.s.l.(Hawksworth and Wiens 1996).
Arceuthobium vaginatum subsp.vaginatum is a dark brown or black(sometimes reddish),hemiparasitic shrub 20–30 cm tall.The dioecious plants have a remarkable sexual dimorphism,with the staminate individuals larger than the pistillate.Anthesis usually occurs in March and April,and fruits reach maturity in August of the next year.Arceuthobium vaginatumparasitizes 13 species of Mexican pines;it is distributed from northern to central Mexico,in an altitudinal range of 2100–3900 m a.s.l.(Hawksworth and Wiens 1996).
The host for these two dwarf mistletoes in the study site is Pinus hartwegii(Pinaceae),which is an evergreen coniferous tree 15–30 m tall,distributed from Mexico to Honduras in temperate subhumid zones at 2850–4200 m a.s.l.It has thick,rough,scaly,dark brown to gray bark;needles 7–14 cm long are in fascicles of 2–6(de Rzedowski et al.2001).
Sampling method
We selected five Pinus hartwegii stands of approximately 100 m2and at least 500 m from each other;the two dwarf mistletoe species were present in each stand.The sites were located between 19°17′0.5″and 19°18′19″N and 98°42′26.4″and 98°40′30.7″W at 3270–3430 m a.s.l.In each stand,we collected stems from both mistletoe species and terminal branches with needles ofP.hartwegiithat were infected by any of the two mistletoe species.Approximately,725 cm3of tissue were collected for each species,from at least five different plants.This procedure was repeated every 2 months for 1 year,from March 2010 to January 2011 for a total of 90 samples:5 sites×3 studied species×6 samplings.
Each sample was processed in a Berlese–Tullgren funnel for 3 days without heat and 3 days with a heat source,after which we collected the arthropods from the bottom of the funnel for posterior counting and identification to the finest taxonomic level possible.Different identification keys were used,according to the relevant taxonomic group.
Data analysis
To evaluate sampling efficiency,we built two species accumulation curves,one for all arthropod species pooled together and one separating the arthropods associated with each plant species(Soberón and Llorente 1993).For the two curves,we implemented the Vegan package of R(R Core Development Team 2015)using the random method with 100 permutations.We also computed species richness(S),abundance(N),diversity(H′),evenness(J′),Jackknife first order(S′,a species pool estimation)and the percentage of reached richness in the samples(based on the Jackknife estimator).The last two were also calculated for total arthropod community.Diversity was estimated with the Shannon–Wiener index(H′)using natural logarithm(Krebs 1999);for evenness,we used the Pielou Index(J′)(Krebs 1999).
Then,we compared species richness similarity between the three plant species with the Jaccard similarity index(SIj)(Krebs 1999).To estimate community similarities from species relative frequencies,we constructed a Bray–Curtis similarity dendrogram(Legendre and Legendre 1998).We also quantified the number of morphospecies present exclusively in one of the three plant species(i.e.,two mistletoes and pine)and the number of morphospecies that were shared among the plant species.These results were illustrated in a Venn diagram.
Community composition to the order level was compared among species and sampling date using a contingency table to examine the frequency of each order with each plant species.The table was tested with a χ2test of independence(Zar 2010),and,if significant,a standardized residues test was performed(Zar 2010).
To determine the effect of the plant species and the sampling month on arthropod species richness,diversity and abundance,we conducted a repeated measure ANOVA for each attribute(Zar 2010).When the latter was signi ficant(p<0.05),we performed the post hoc Tukey test to identify homogeneous groups.In cases where the Tukey test did not show any significantly different groups(even when the ANOVA test was significant),we used the Fisher test(Zar 2010).
To determine differences among the arthropod community assemblage of three habitat types(pine,A.vaginatum and A.globosum),we used a principal coordinate analysis(PCoA).The Adonis test was used for finding significant differences in abundance of taxa comparing pine versusA.vaginatumversusA.globosumtreatments of arthropods communities.When the Adonis test was significant(p<0.05),additional Adonis tests were done between all possible pair comparisons of treatments(pine vs.A.vaginatum,pine vs.A.globosum,A.vaginatum vs.A.globosum).All statistical analyses were done in the Vegan package of R(R Core Development Team 2015).
Results
General community composition
A total of 32,059 individuals were collected(both adult and immature stages)from 89 samples(A.vaginatum sample from March went missing).Of these,51 morphospecies were recognized;20,619 individual were collected during the rainy season(May,July and September)and 11,440 during the dry season(January,March and November).Of the 51 morphospecies,15 were identified to order level,26 to family,six to genera and only four to species(see Appendix for details).
Fifteen orders were determined,listed from most to least abundant:Prostigmata,Thysanoptera,Oribatida,Homoptera,Collembola,Psocoptera,Hymenoptera,Hemiptera,Coleoptera,Mesostigmata,Lepidoptera,Diptera,Araneae,Opiliones and Mecoptera(orders according Borror et al.1989;Fig.1).Subclass Acari was very abundant on the three speciesand wasrepresented by three orders:Prostigmata,Mesostigmata,and Oribatida.Prostigmata was the dominant group in the three plants,and the most abundant genus was Brevipalpus.For Oribatida,the greatest abundance and diversity were associated with P.hartwegii,on which 8%of the specimens were found.Mesostigmata,however,was very infrequent(<2%),and most individuals belonged to the family Phytoseiidae.
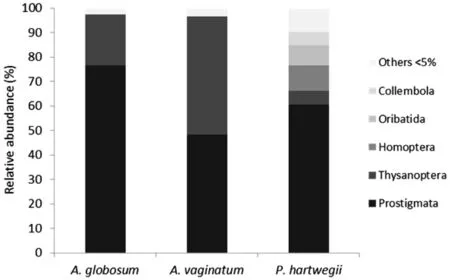
Fig.1 Relative abundance(%)of the arthropod orders associated with P.hartwegii(N=4294),A.globosum(N=12,224)and A.vaginatum(N=15,541).“Others”are orders with<5%of abundance(Psocoptera, Hymenoptera,Hemiptera,Coleoptera,Mesostigmata,Lepidoptera,Diptera,Araneae,Opiliones and Mecoptera)
Thysanoptera,the second most abundant order,represented 21,48 and 5%of the arthropods associated withA.globosum,A.vaginatum and P.hartwegii,respectively(Fig.1).At least three species were found:Frankliniella tolucensis,F.fallaciosa and F.minuta,plus a possible new species for the genus.Although Homoptera had a relative low abundance,it represented 10%of the arthropods associated withP.hartwegii.Collembola order,represented by 21 species of suborder Entomobryomorpha and 22 morphospecies of suborder Poduromorpha,represented 5%of the abundance on pine needles,while on both mistletoes it contributed<1%of the abundance.The remaining nine orders(Psocoptera,Hymenoptera,Hemiptera, Coleoptera,Lepidoptera,Diptera,Araneae,Opiliones and Mecoptera)represented<5%of the abundance for each plant species.
Sampling efficiency was high because the arthropod richnesses were very close to the species pool estimations(S′)for the total community(Fig.2a)and for each plant(Fig.2b),and due to the elevated percentages of the richnesses(>78%;Table 1).
The Bray–Curtis dendogram and Jaccard similarity index(SIj)showed that similarity between communities is greater between both dwarf mistletoe species(SIj=0.625),than any of the mistletoes with P.hartwegii(A.vaginatum SIj=0.53;A.globosum SIj=0.59;Fig.3).Likewise,most of morphospecies(47.05%)were present in the three plants,and a few were exclusive for one of the plants(A.vaginatum=11.76%,A.globosum=5.88%,P.hartwegii=19.60%;Fig.4).
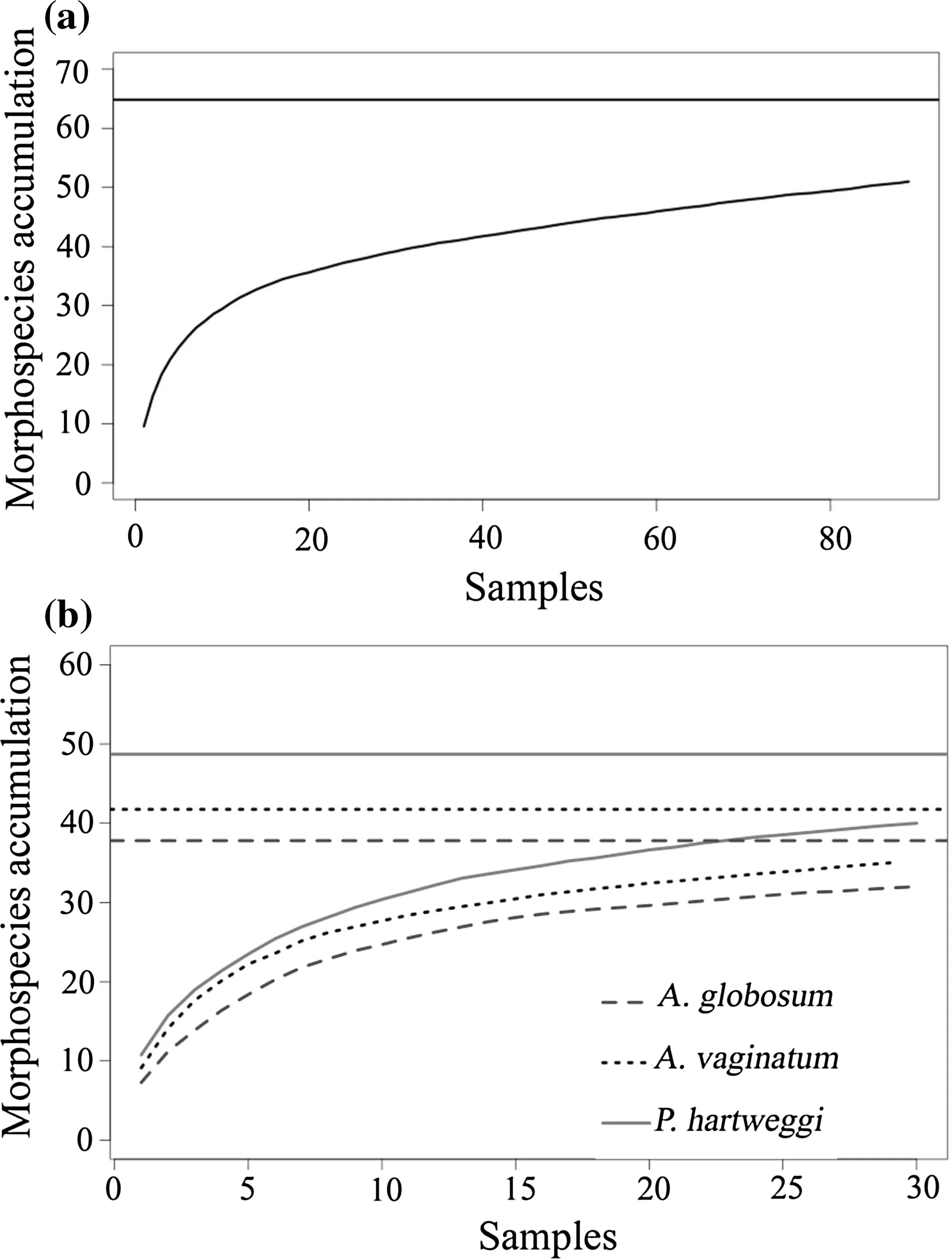
Fig.2 Accumulation curve of a pooled morphospecies and b morphospecies for each plant species.Horizontal lines represent species pool estimation(S′)for each plant
Community composition per plant species
Arceuthobium vaginatumandA.globosumhad the greatest values of abundance and density of arthropods,but P.hartwegii had the greatest values of species richness,diversity and evenness(Table 1).
The abundance of the most abundant arthropods(Prostigmata and Thysanoptera)depended on the type of plant(χ2=9539,df=28,p<0.001;Fig.5a).Prostigmata species were significantly more abundant inA.globosumthan expected from random,and in the same way,significantly lower inP.hartwegii(Fig.5a).Meanwhile,individuals of Thysanoptera were significantly more abundant in A.vaginatum and significantly lower in A.globosum(Fig.5a).
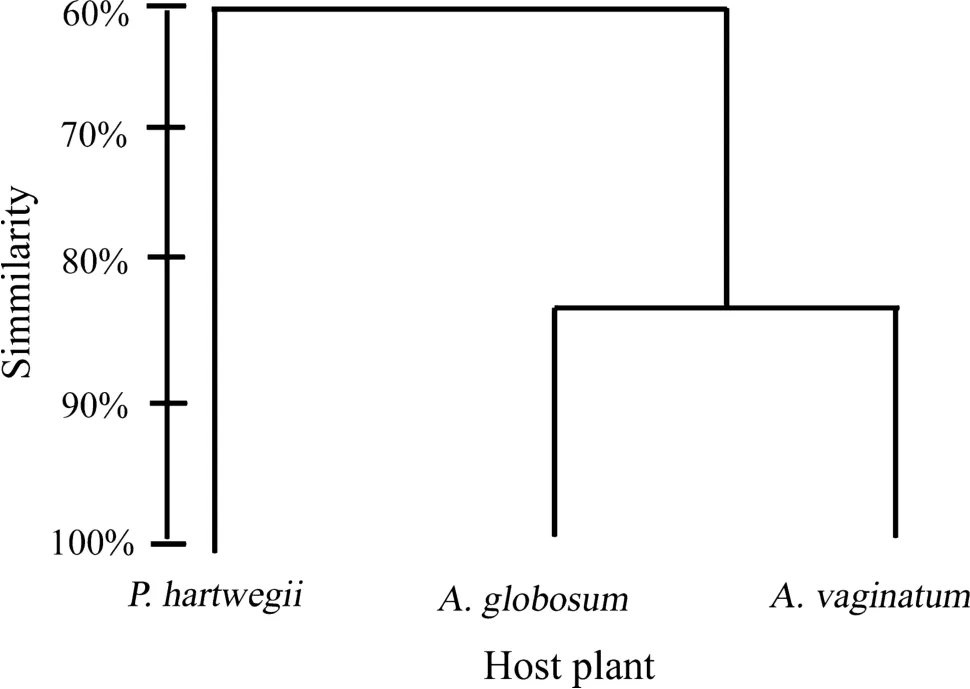
Fig.3 Bray–Curtis dendogram comparing arthropod communities associated with A.globosum,A.vaginatum and P.hartwegii
The abundance of each order was also dependent on sampling month(χ2=14,625,df=55,p<0.001;Fig.5b).The most notable trends were for Prostigmata,which was significantly more abundant in July,September,November and January(than expected for random),and for Thysanoptera,which was significant more abundant in March and May(Fig.5b);on the contrary,the frequencies of Prostigmata and Thysanoptera were significant lower in the remaining months of sampling,for each case(i.e.,March and May for Prostigmata,and July,September,November and January for Thysanoptera;Fig.5b).
Effect of plant species and sampling month on the community attributes
There were significant effects of the plant(F2,14=14.91,p<0.001),the sampling month(F5,14=4.02,p=0.003),and their statistical interaction(plant×sampling month;F10,14=2.90,p=0.046)on the arthropod morphospecies richness.Arthropod richness was higher onP.hartwegiiandA.globosumcommunities than onA.vaginatum(Fig.6a).Significant differences in sampling month were only found in March and May;P.hartwegii had a higherrichness in March than the other two plants did,whereas A.globosumhad a lower richness on May(Fig.6b).
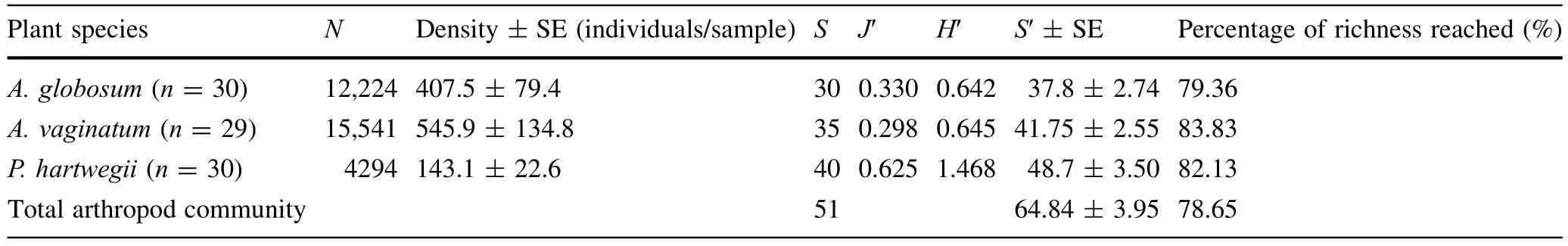
Table 1 Abundance(N),density,morphospecies richness(S),evenness(J′),diversity(H′),Jackknife first order(S′)and percentage of reached richness(using Jackknife species pool estimator)of arthropods associated with three plant species
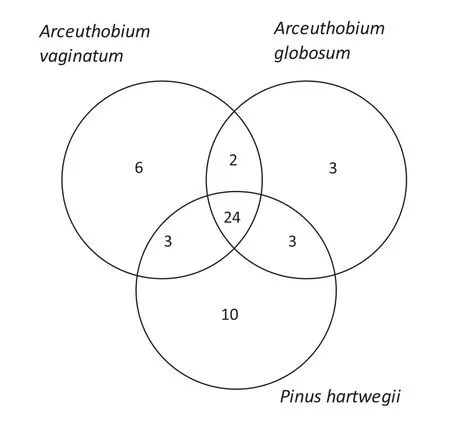
Fig.4 Number of arthropod morphospecies exclusive to and shared by the three plant species studied in this work(A.vaginatum,A.globosum or P.hartwegii)
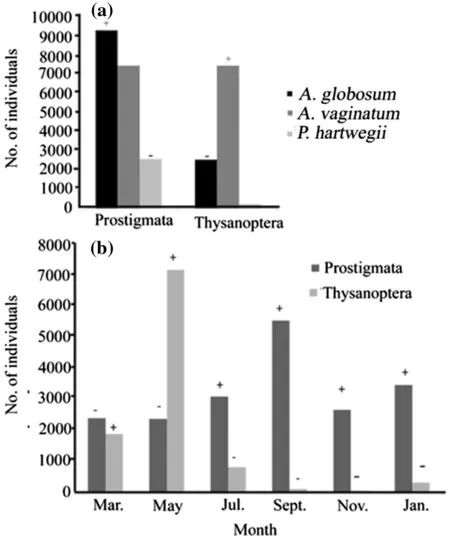
Fig.5 Absolute frequencies of the most abundant orders of arthropods a on each plant and b for each sampling month.+higher frequency than expected;−lower frequency than expected
We also found a significant effect of plant species(F2,14=14.9,p<0.001),sampling month(F5,14=4.5,p=0.001),and their interaction(plant×sampling month;F10,14=3.6,p=0.001)on arthropod mean abundance.Arthropod abundance was significantly higher on the two mistletoe species,than on the pine host(Fig.7a).On the other hand,mean abundance on A.vaginatum was significantly higher only in May than the other two plant species(Fig.7b).

Fig.6 a Morphospecies richness(±SE)on each plant species.b Morphospecies richness(±SE)on each plant species by sampling month.Letters represent significant differences among groups,according to Tukey’s post hoc test(p<0.05)
For diversity(H′),there was a significant effect of plant species(F2,14=47.47,p<0.001)and sampling month(F5,14=2.57,p=0.037),but not for their interaction(F10,14=1.22,p=0.299).The community associated with P.hartwegii was more diverse than the communities associated with the mistletoes(Fig.8a).Moreover,diversity was higher in March,declined over time,and diversity in March was higher than in November and January(Fig.8b).
According to the PCoA,the arthropod communities differed significantly among the three habitat types(Adonis test,p<0.001;Fig.9),besides there were significant differences between arthropod communities of pine versus A.vaginatum(Adonis test,p<0.001)and pine versus A.globosum(Adonis test,p<0.001);nevertheless,A.vaginatum versus A.globosum comparison did not show any significant difference(Adonis test,p=0.30;Fig.9).
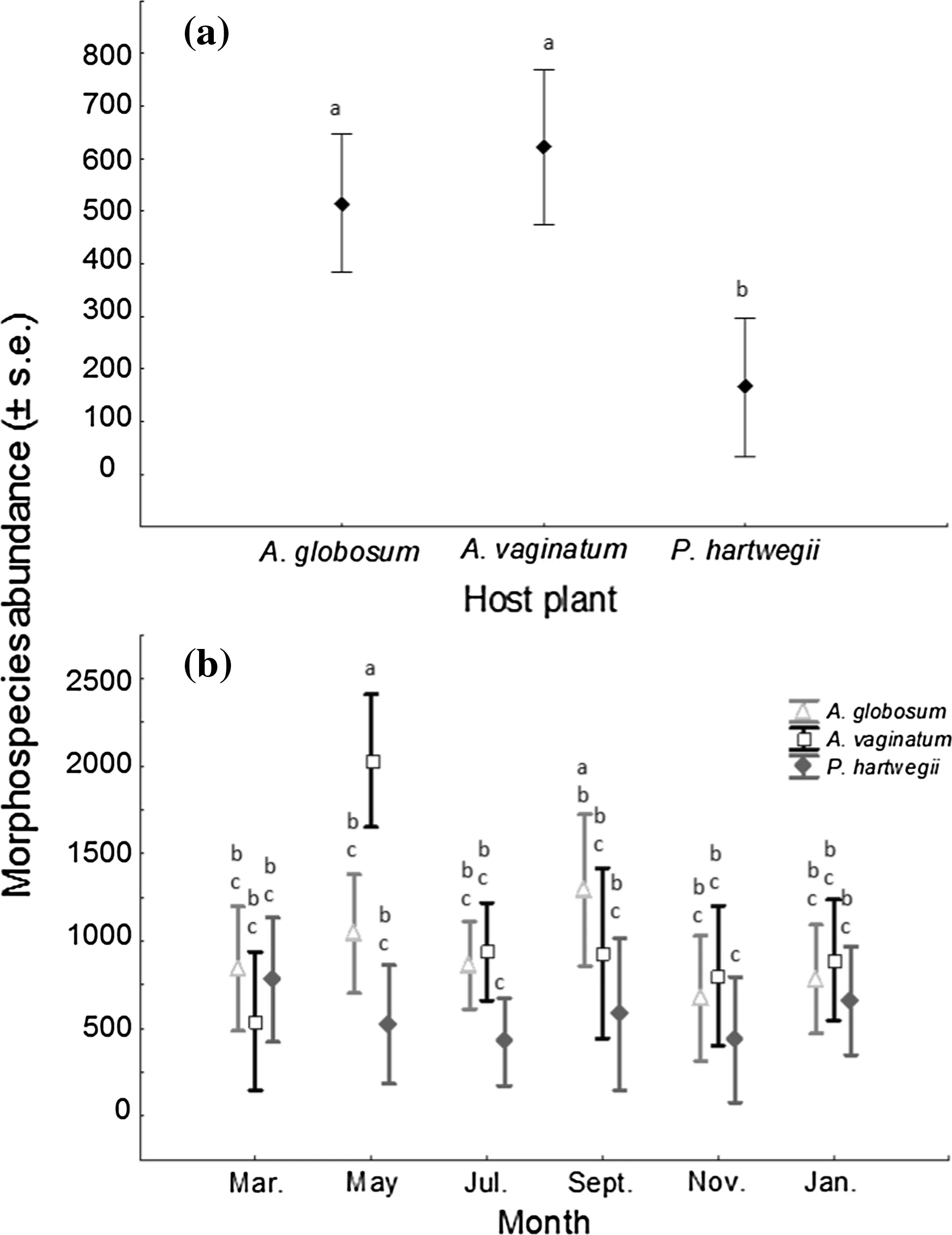
Fig.7 a Morphospecies abundance(± SE)on each species.b Morphospecies abundance(±SE)on each species by sampling month.Letters represent significant differences among groups,according to Tukey’s post hoc test(p<0.05)
Discussion
Community structure and temporal variation
Results clearly showed thatP.hartwegiihas greater morphospecies richness,diversity and evenness,and a lower abundance of arthropods compared with dwarf mistletoes’aerial tissues.Communities associated with the two mistletoes have similar abundances of certain groups,such as Thysanoptera and Prostigmata,which can be attributed to their phylogenetic closeness and great phenotypic resemblance(Hawksworth and Wiens 1996),thus the provision of similar resources and conditions.

Fig.8 a Diversity(H′)on each species and b in each sampling month.Letters represent significant differences among groups,according to Tukey’s post hoc test(p<0.05)
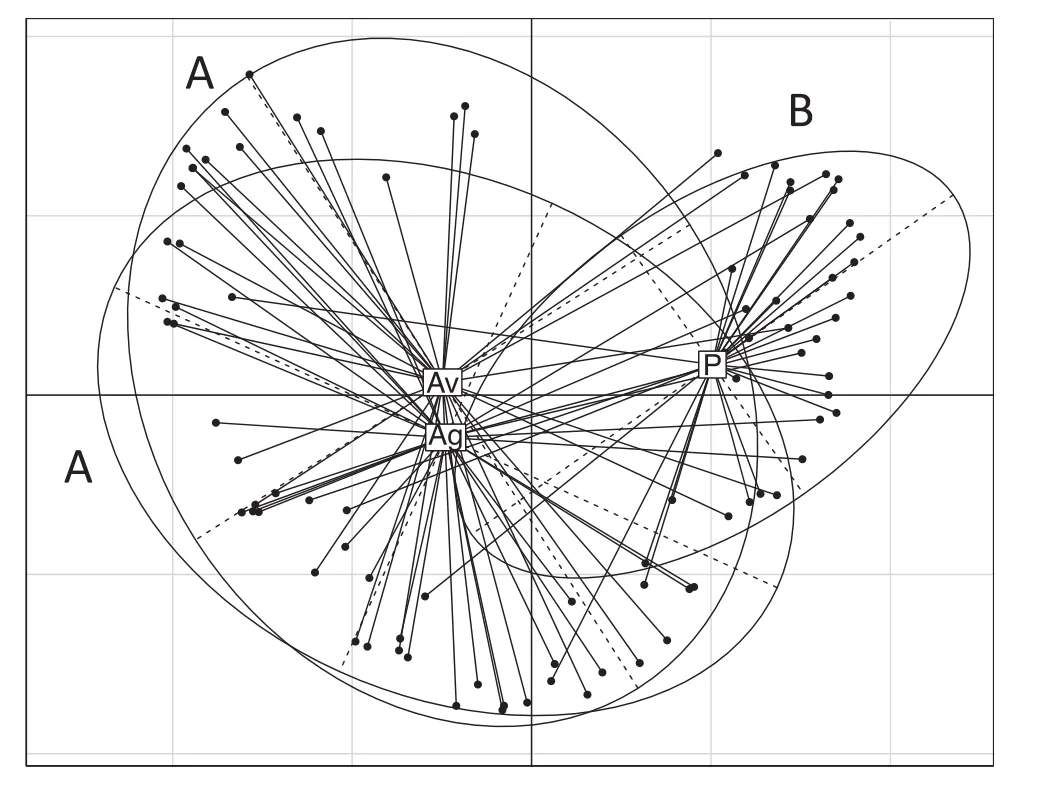
Fig.9 PCoA analysis comparing species abundance in the arthropod community samples from the three habitats:pine(P),A.vaginatum(Av),A.globosum(Ag).Different letters denote significant differences in an Adonis test(p<0.05)
The great diversity of arthropods found onP.hartwegiineedles can be attributed to two things:(1)this species provides a larger colonization island than the mistletoes do(Schoener 1976;Connor and McCoy 1979;Lomolino 2000;Williamson et al.2001;López-Gómez and Cano-Santana 2010),and(2)needles on terminal branches have a rosette-like shape,sheltering arthropods from climatic variations and predators(López-Gómez and Cano-Santana 2010).The scale-like leaves of the dwarf mistletoes provide a smaller colonization island and are ineffective shelters for many arthropods,especially those of medium to large size.In addition,the higher diversity of arthropods on pine needles is likely favored by tourist species(i.e.,a nonpredatory species that does not have a lasting association with a host plant;Moran and Southwood 1982)looking for the more heterogeneous resources that pines provide.Individuals belonging to Homoptera,Coleoptera,Diptera,Hemiptera,Homoptera,Araneae and Hymenoptera were found living inside pine fascicles,which also tend to accumulate debris(our personal observations)that can be used as feeding sites by saprophagous arthropods such as springtails,oribatid mites,Opiliones,and Psocoptera bark flies(Borror et al.1989;Schowalter 2011).
The greater variety of habitats and resources of the pine foliage may also increase the number of trophic levels that favor top predators such as spiders,mites and parasitic wasps Mesostigmata(Schoener 1989;Post et al.2000).Winged insects,however,may be underestimated since they can fly to avoid capture.
Although A.globosum(up to 70 cm tall)and A.vaginatum(maximum height 55 cm)clearly differ in size(Hawksworth and Wiens 1996),our results showed that the smallerA.vaginatumhad a greater abundance,density and richness of arthropods than didA.globosum.However,we collected the same volume of tissue for both species,regardless of their size,so we cannot draw precise conclusions about any size effect.Moreover,in a study in 2009(a year before this study)in the same area,A.vaginatum was more prevalent than A.globosum(27.6 and 6.7%of infected trees,respectively),as well more abundant(mean dwarf mistletoe rating system:2.05 and 1.18,respectively)(Queijeiro-Bolaños et al.2014).Thus,despite its sizeA.vaginatumapparently provides more available habitat for the arthropod community.Further studies are clearly necessary to clarify contributions of size and abundance to the arthropod community composition.
Arthropod communities on dwarf mistletoes had a lower species richness compared with that reported for other mistletoe species such as Decaisnina signata(113 arthropod species and 12 orders;Anderson and Braby 2009)andAmyema miquelli(10 orders;Burns et al.2011).Loranthaceous mistletoes are generally larger(including branches,leaves, flowers and fruits)than dwarf mistletoes and may thus offer greater colonization space and feeding resources.In contrast,dwarf mistletoes have small scalelike leaves that may be a more suitable refuge for microarthropods(body length<5 mm)such as thrips and mites.Because of their smaller flowers,dwarf mistletoes may depend on insects such as encyrtid wasps(Hymenoptra:Encrytidae)for pollination(when they are not wind-pollinated)(Pen field et al.1976).
Arceuthobium vaginatum presents a marked sexual dimorphism,with staminate plants usually larger than the pistillate(Hawksworth et al.2002),but A.globosum does not.We did not differentiate between male and female plants,and as far as we know,no biotic associations have been correlated with the sexual dimorphism of mistletoes.However,differences in species richness and composition can be expected based on variations in size derived from sexual dimorphism;larger mistletoes can host greater species richness(Anderson and Braby 2009).It has been reported for plants in general that,depending on the gender,nutrients can vary,and frequently arthropods prefer male plants(Agren et al.1999).Nevertheless,studies focused on sexual dimorphism in mistletoes are still needed.
Temporal variation of arthropod communities showed subtle changes in richness and composition;the greatest abundance of organisms was present in the rainy season(May–October),with a notable increase in abundance of individuals belonging to orders Thysanoptera,Collembola,Psocoptera, Hymenoptera,Coleoptera and Oribatida.Throughout this study,we did not record humidity or leaf production or herbivory rates because the rainy season is known to be strongly related to increased presence of herbivores as a direct consequence of greater resource availability(Basset 1996;Stork et al.1997).
Functional role of arthropods
We found 15 orders of arthropods,which include phytophagous and predatory organisms,on the three plant species.Stevens and Hawksworth(1970,1984)described generalist and specialist insects feeding or living within dwarf mistletoes;the most frequent belonged to the orders Lepidoptera, Hemiptera, Coleoptera, Thysanoptera, Prostigmata,Mesostigmata,and Araneae.Even so,there is very little evidence that phytophagous insects on dwarf mistletoes in Mexico can function as agents to control these parasitic plants.
Prostigmata was the most abundant order of arthropods associated with the three plant species;the species within these order are reported as phytophagous(Borror et al.1989),and some have been described as specialist shoot predators for dwarf mistletoes(such asTyphlodromusarceuthobius[Phytoseiidae],T.pusillum,Paraphytopus arceuthobii [Eriophyidae] and Brevipalpus porca[Tenuipalpidae];Keifer 1952;Pritchard and Baker 1958;Kennett 1963).Although we do not know the identity of all the morphospecies,the genus Brevipalpus and members of the families Phytoseiidae were present,indicating that they could act as predators for dwarf mistletoes’shoots.Mites are also a major group of saprophagous insects(Maraun and Scheu 2000)that can carry out the facilitation process,decomposition and nutrient cycling(Visser 1985).
On the other hand,the presence of Collembola in forest canopies has been reported as largely due to the association they have with epiphytic plants(Prinzing 1997).Although springtails are considered general consumers,they can feed on fungal hyphae and spores,bacteria,detritus,pollen and mineral particles,and some have predatory habits(Castaño-Meneses et al.2004).The rosette-like structure of pine needle fascicles may increase the retention of debris,favoring the presence of saprophagous arthropods such as springtails.
Thysanoptera was the second most abundant order.Thrips are commonly associated with dwarf mistletoes and have been described as shoot feeders(Stevens and Hawksworth 1970,1984).The highest abundance of thrips(especiallyFrankliniella tolucensis)was reported in the May sample ofA.vaginatum,when it was flowering(Hawksworth and Wiens 1996).Thus,thrips could be pollinivores,although this possibility needs confirmation.These thrips are also an important prey for Phytoseiidae mites and egg feeding wasps(Morse and Hoddle 2006).
Although the homopteran members could not be identified to a more specific taxonomic level,they probably belong to the Mitiidae family,which are shoot feeders ofA.vaginatumsubsp.cryptopodumin the southern United States and northern Mexico(Kuijt 1955).
Adult Curculionidae beetles(Coleoptera)often oviposit on pine branches,and the hatching larvae feed on pine buds(Hawksworth and Wiens 1996).So beetle larvae might also exploit mistletoe shoots as a feeding resource.
It is feasible that wasps(Hymenoptera)are closely associated with the three plant species.Some wasp species feed on nectar and pollen of dwarf mistletoe flowers(Goulet and Huber 1993),but they can parasitize other arthropods,such as members of Araneae,Coleoptera,Heteroptera,Homoptera and Lepidoptera,which were all present in the three plant species.
Members of the order Lepidoptera,such as the phytophagous larvae of Mitoura sp.and Neophasia sp.found on A.vaginatum subps.cryptopodum(Grimble and Beckworth 1993)may be mistletoes specialist consumers.We therefore need to determine the morphospecies of the lepidopterans to the finest taxonomic level(which we were not able to do,since we found only larvae)because these specialist organisms may have potential to control dwarf mistletoe.
Spiders are also predators and likely to be able to feed on all the other arthropods present on the mistletoes and the pine;it also can be parasitized by wasps.
Conclusions
Each of the three plant species maintain different arthropod communities,which are very similar to each other,possibly because of the phylogenetic proximity of the three species.The pine species,however,is able to host a greater variety of organisms by providing a wider range of resources and refuge sites.
The role that parasitic plants such as mistletoes play in their communities can be diverse and complex,as was evident in our study system.Although dwarf mistletoes have been traditionally viewed as harmful parasites,they can also be a positive component of the canopy by supporting a vast arthropod community and providing suitable microhabitats and resources for a great variety of organisms.The great abundance of microarthropods found within these parasitic plants is supportive evidence for the theory that mistletoes act as a key group within their biological communities(Watson 2001;Watson and Herring 2012).
Deeper studies on the phytophagous groups(Thysanoptera,Prostigmata and Lepidoptera)should focus on their potential as biocontrol agents for dwarf mistletoe.
The study of such complex systems(like pine-mistletoearthropods)in forests should lead to more integrative strategies for forest management.
AcknowledgementsWe thank Iván Castellanos-Vargas for his technical support and Arturo Garcıá for critically reviewing this work.Invaluable support for identifying specimens was provided by Ricardo IglesiasMendoza(Cryptostigmata),Fernando Álvarez Padilla(Araneomorphae), Roberto Johansen Naime and Elizabeth Mejorada(Thysanoptera)and Javier Garcıá Figueroa(Psocoptera).This study was supported by UNAM-DGAPA-PAPIIT IN220912 Grant to ZC-S.
Appendix
List and abundances of arthropods species associated with the dwarf mistletoes Arceuthobium globosum and A.vaginatum,and their host Pinus hartwegii.Organisms were collected in the Iztaccíhuatl-Popocatépetl National Park,Mexico.The taxonomic classification was based on Borror et al.(1989).

Morphospecies Taxonomic classification Host plant A.globosum A.vaginatum P.hartwegii Phylum Artrhopoda Class Arachnida Order Araneae Suborder Araneomorphae Superfamily Araneoidea Family Araneidae 1 Araneidae sp.1 1 0 0 2 Araneidae sp.2 0 1 0 Family Theridiidae 3 Theridiidae sp.1 7 5 2 4 Theridiidae sp.2 0 1 0 5 Theridiidae sp.3 0 0 1 Family Linyphiidae 6 Linyphiidae sp.1 0 3 0 7 Linyphiidae sp.2 0 0 1 Family Tetragnathide 8 Tetragnathide sp.1 0 0 3 Order Opiliones 9 Opilionide sp.1 0 1 1 Class Acari Order Mesostigmata Suborder Monogynapsida Cohort Gamasina Subcohort Dermanyssiae Super Family Phytoseioidea Family Phytoseiidae 10 Phytoseiidae sp.1 5 16 10 11 Phytoseiidae sp.2 5 5 48 12 Phytoseiidae sp.3 5 0 3 Suborder Prostigmata Cohort Raphignathina Superfamily Tetranychoidea Family Tenuipalpidae Genera Brevipalpus 13 Brevipalpus sp.1 9304 7427 2400 Superfamily Tetranychoidea Family Tetranychidae 14 Tetranychidae sp.1 15 45 153 Superfamily Raphignathoidea Family Camerobiidae Genera Camerobia 15 Camerobia sp.1 18 8 12 Cohort Parasitengomina Superfamily Erythraeoidea Family Erythraeidae 16 Erythraeidae sp.1 8 12 36 Suborder Oribatida Cohort Brachypylina

Table a continued
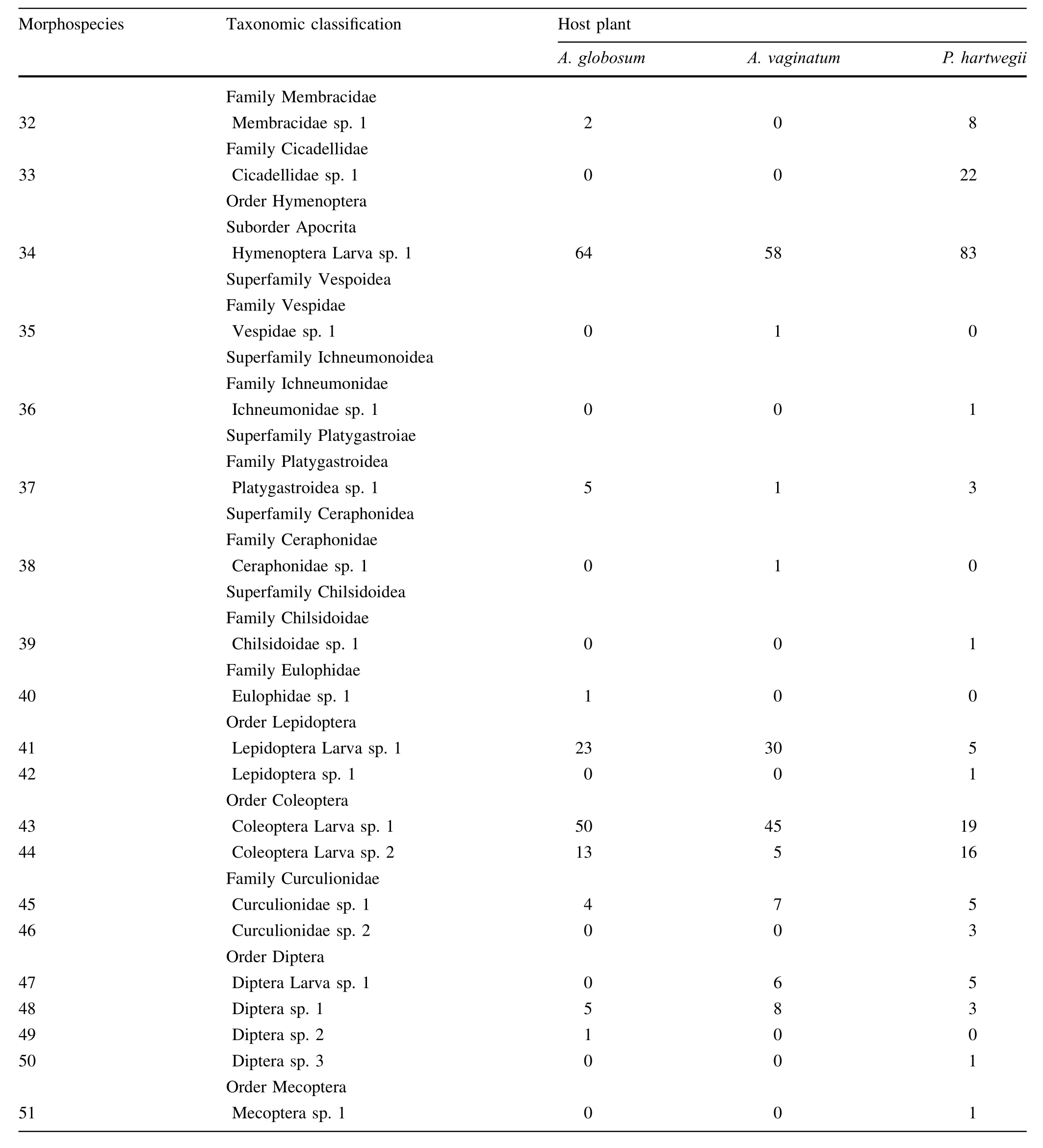
Table a continued
 Journal of Forestry Research2018年5期
Journal of Forestry Research2018年5期
- Journal of Forestry Research的其它文章
- Environmental load of solid wood floor production from larch grown at different planting densities based on a life cycle assessment
- An overview of proven Climate Change Vulnerability Assessment tools for forests and forest-dependent communities across the globe:a literature analysis
- Assessing the vulnerability of a forest ecosystem to climate change and variability in the western Mediterranean sub-region of Turkey:future evaluation
- Effects of management regimes on carbon sequestration under the Natural Forest Protection Program in northeast China
- Mixed-effects modeling for tree height prediction models of Oriental beech in the Hyrcanian forests
- Estimation of a basal area growth model for individual trees in uneven-aged Caspian mixed species forests