
两种叶型楠木叶片表观特征及光合特性的比较分析
2018-08-29 07:37吴小林姚月华蒋仲龙杨平昆张茂付童再康
浙江林业科技 2018年3期
吴小林,姚月华,方 霞,蒋仲龙,杨平昆,张茂付,童再康
两种叶型楠木叶片表观特征及光合特性的比较分析
吴小林1,姚月华2,方 霞3,蒋仲龙4,杨平昆1,张茂付1,童再康3
(1. 浙江省庆元县实验林场,浙江 庆元 323800;2. 浙江省庆元县林业局,浙江 庆元 323800;3. 浙江农林大学 亚热带森林培育国家重点实验室,浙江 杭州 311300;4. 浙江省林业厅,浙江 杭州 310020)
为研究大叶楠木f.和小叶楠木f.两种叶型楠木幼树叶片表观特性和光合特性差异及其影响因子,本文以2年生两种叶型的楠木为研究对象,测定叶片形态指标和夏季光合特性参数,分析影响净光合速率的相关因子。结果表明:(1)两种叶型楠木的叶面积、叶片长、叶片宽和叶脉数存在显著差异(<0.05),不同冠层的叶片也存在显著差异;(2)大叶楠木的净光合速率显著高于小叶楠木,由多元逐步回归分析与通径分析显示,影响大叶楠木净光合速率的主要因子是气孔导度、胞间CO2浓度、叶片水分亏压、叶温和蒸腾速率5个因子,其中气孔导度起决定作用。而影响小叶楠木净光合速率是叶片水分亏压、空气相对湿度、大气二氧化碳浓度、蒸腾速率和胞间CO2浓度5个因子,且叶片水分亏压对净光合速率影响最大。两种叶型楠木幼树在夏季需要适当遮荫和充足的水分供应,以增加光合速率与有机物积累量。
楠木;叶型;表观特性;光合特性;净光合速率
光合作用是植物生长和代谢活动的生理基础[1-2]。同一物种不同类型间、品种间的光合效率存在显著差异,郑彩霞等研究2年生盆栽胡杨锯齿卵圆形、卵圆形和披针形多形叶的形态、解剖和光合特性,叶片在发育过程中为适应干旱荒漠的生态环境发生变异[3]。王海珍等研究新疆塔里木河上游人工林胡杨条形、卵形和锯齿阔卵形叶,比较其光合作用-光与CO2响应及叶绿素荧光响应特征[4]。褚延广等测定108个欧洲黑杨无性系的气体交换参数、叶绿素荧光及生长参数[5]。黄红英等研究了贵州和海南两种生态型麻疯树L夏天气体交换参数和叶绿素荧光参数[6]。前人研究发现同一物种不同类型间的光合效率与自然生境和品种密切相关,但夏季同一物种不同类型间的净光合速率日变化均呈双峰曲线,出现了光午休现象,如红砂Linn[7],蔷薇[8],梨spp[9],葡萄[10]和天山雪莲[11]等植物。叶片是植物光合作用的主要器官,影响树木生长量和有机物积累量[12-13]。因此研究同一物种不同叶型的的叶表观特征与光合特性,对于选育高光效植物品系具有重要意义。
楠木属于樟科Lauraceae楠属常绿大乔木,树干通直,树姿优美,材性优良,木材纹理顺直,耐腐防蛀,是木结构建筑的最佳用材树种,是制作高级家具的上等用材,也被广泛运用于绿化景观营造。楠木的研究主要集中在抗逆性、群落特性和光合特性等方面。Akash Tariq等研究指出在干旱胁迫下施加磷肥可以提高楠木幼苗的抗旱性[14]。Hu Y等发现楠木幼树在轻度和中度干旱胁迫后,复水能恢复[15]。李高志等发现在低温胁迫下1年生大叶楠木苗的抗寒能力强于小叶楠木[16];宗卫等发现自然低温下水杨酸(1 mmol·L-1)处理可以提高1年生闽楠和楠木幼苗植株的耐寒性[17]。Gao J等利用AFLP分子标记技术研究中国6个省份楠木种群结构和遗传多样性[18]。王茹等研究不同光环境下楠木叶绿素荧光特性[19]。胡婧楠等研究2年生楠木和华东楠光合生理特性[20],殷国兰等研究1年生樟,楠木和红椿的光合生理特性[21]。然而,有关楠木种内变异及其光合性能差异等方面的研究还未见报道。本项研究以来自相同产地的2年生大叶楠木和小叶楠木两种叶型楠木为研究对象,测定叶表观特性和光合速率日变化,探寻两种叶型楠木的叶片及其光合特性差异,为楠木的栽培和优良品系选育提供科学依据。
1 材料与方法
1.1 试验地概况
试验设于浙江省庆元县实验林场,地理坐标118°50′ ~ 119°30′ E,27°15′ ~ 27°51′ N,属亚热带季风区,温暖湿润,四季分明,年平均气温17.4℃,年降水量1 760 mm,无霜期245 d。
1.2 试验材料
大叶楠木和小叶楠木的试验种子采自四川省雅安荥经县,种子于2015年1月育苗,基质为黄土:树皮:谷壳:珍珠岩=4:3:2:1,基质中加施缓释肥3 kg·m-3,自动灌溉水分管理,幼苗阶段适时遮荫50%。试验材料为2 年生容器苗,试验随机选取大叶楠木和小叶楠木各100株。
1.3 研究方法
1.3.1 叶表观特性测定 将楠木植株分为上中下3层,各层均选取同一部位的3片叶用于测定形态指标。随机选取100株取叶样,取样叶片连带叶柄。用叶面积扫描仪YMJ-B扫描获得叶片长、叶片宽、叶面积和长宽比等指标。用游标卡尺测量叶柄长,计数叶脉数,测定苗高、地径。
1.3.2 气体交换参数日变化测定 于2017年7月,选择晴朗天气,采用Li-6400便携式光合测定系统,选取生长良好植株的冠层中上部正常叶片,无遮光,让其处于自然生长状态,从8:00-18:00,每隔2 h测定1次,各选取6株,每株测定3片叶。
1.4 数据处理
使用Microsoft Excel 2010软件进行数据处理和表格制作,SPSS 19.0软件进行方差和相关性分析,采用Graghpad Prism 5绘图。
2 结果与分析
2.1 两种叶型楠木的叶表观特征
经单因素方差分析,发现2年生同种叶型楠木不同层次间的叶面积、长度、宽度和长宽比、叶柄长和叶脉数均存在显著差异(<0.05)(见表1)。楠木以中层叶片均值最大(1 869.39±584.62 mm2),是下层叶片(1 269.52±329.69 mm2)的1.47倍;而小叶楠木则以上层叶片最大(1 576.00±284.54 mm2),是下层叶片(698.15±178.53 mm2)的2.26倍。同一层叶片两种叶型楠木相差168.04 ~ 571.37 mm2,可见两种叶型楠木的各层叶片大小均有明显差异。同时,大叶楠木的叶长、叶宽以及叶脉数均明显高于小叶楠木。
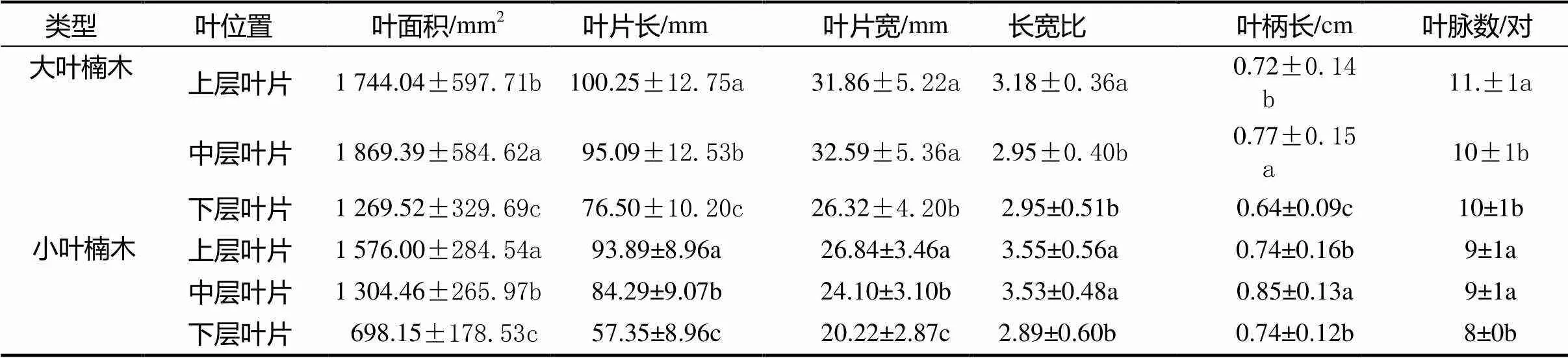
表1 两种叶型楠木叶片表型均值
注:数据以均值标准差表示,同列不同字母表示差异显著(<0.05)。
2.2 两种叶型楠木叶表型性状方差分析
两种叶型楠木表型性状方差分析结果列于表2。可见,叶面积、叶脉数量与叶柄长度3个性状两种类型间、叶片着生部位间以及二者的交互作用均具有极显著(<0.01)的差异。可以认为,生长在同一区域大叶楠木和小叶楠木在叶片表观性状上具有极显著(<0.01)的差异,可以区分两种类型。另外,同一叶型的楠木的不同着生位置的叶面积、叶柄长和叶脉数也具有不同,结合表1可以看出,叶片大小差异最为显著,叶脉数量与叶柄长度相对差异较小。分层测定叶片表型性状进行类型间的比较具有科学性。
表2 两种叶型楠木叶片表观特征性状方差分析
Table 2 ANOVA on phenotypic properties of leaves between two species

表3 同一种楠木的不同层次间的叶片表型指标方差分析
Table 3 ANOVA on phenotypic properties of leaves at different layers of the same species
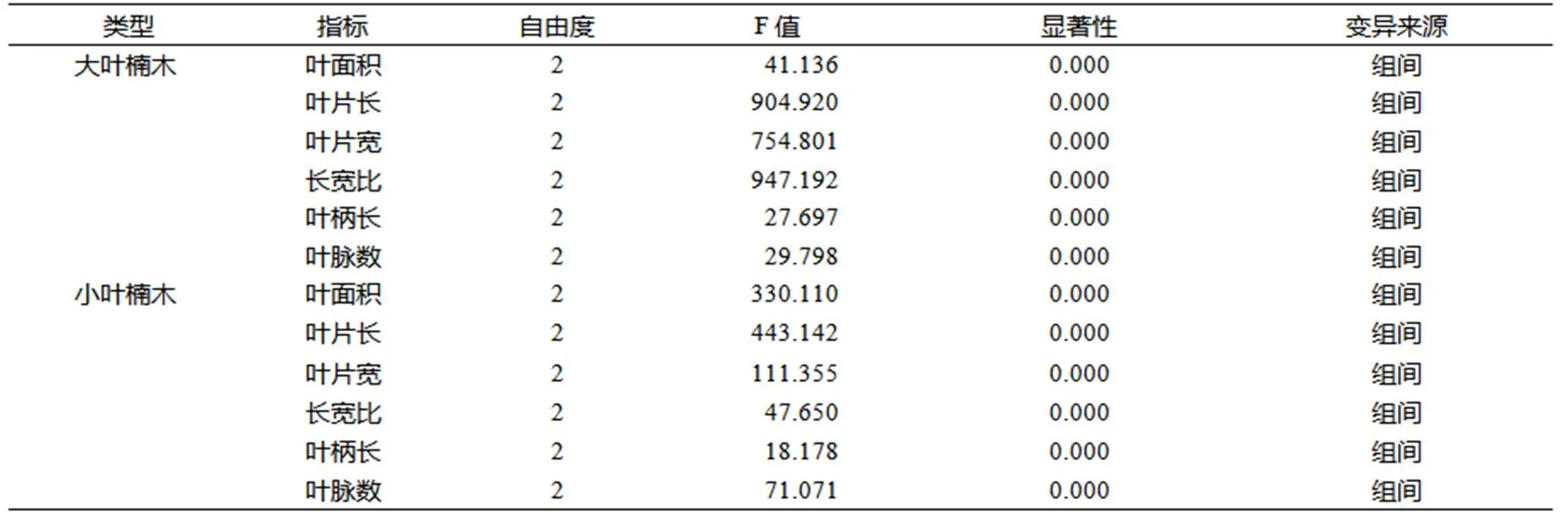
同一种楠木不同层次间的叶片表型指标方差分析结果列于表3。大叶楠木和小叶楠木不同层次间的叶面积、叶片长、叶片宽、长宽比、叶柄长和叶脉数均具有极显著(<0.01)的差异。
2.3 叶片表型特征相关分析
经相关分析知(表4),两种叶型楠木的叶面积、叶片长、叶片宽、叶柄长和叶脉数4个性状间存在极显著(<0.01)正相关,即叶柄长度与叶脉数量均与叶片大小成正比,而且叶片长度与宽度也成正比。说明两种类型楠木的叶片很均匀,主要表现在叶片的大小及厚薄差异上(小叶楠木叶片明显薄)。
表4 两种叶型楠木叶片表型指标相关分析
Table 4 Correlation analysis on leaf phenotypic properties of two species

2.4 两种叶型楠木的光合作用参数比较
表5列出了两种叶型楠木的5个光合参数均值。可见,大叶楠木的净光合速率、气孔导度和光能利用效率显著性高于小叶楠木,其中大叶楠木的净光合速率是小叶楠木1.58倍,气孔导度(0.059 mol·m-2·s-1)也远大于小叶楠木(0.036 mol·m-2·s-1),光能利用率是小叶楠木的3倍;大叶楠木的蒸腾速率和胞间二氧化碳浓度略高于小叶楠木,但差异不显著(>0.05)。
表5 两种叶型楠木的光合作用参数
Table 5 Photosyntheticparameters of two species

注:数据以均值标准差表示,同列不同小写字母表示差异显著(<0.05)。
2.5 光合速率的影响因子分析
为定量分析影响楠木净光合速率Pn的因素,采用多元逐步回归建立模型,以环境因子光合有效辐射PAR(1)、大气CO2浓度Ca(2)、叶温Tl(3)、大气温度Ta(4)、空气相对湿度RH(5)和光合作用其它参数如气孔导度(6)、胞间CO2浓度(7)、蒸腾速率(8)、水分利用效率(9)、叶片水分亏压(10)与净光合速率()进行多元逐步回归。
大叶楠木的多元逐步回归方程:
该模型筛选出气孔导度、胞间CO2浓度、叶片水分亏缺、叶温和蒸腾速率5个显著的影响因子,说明大叶楠木的净光合速率主要受这5个因子的影响。为了进一步确定各影响因素的作用大小,进行通径分析(表6)。从直接作用绝对值看,在夏季晴天,各因子对净光合速率的影响顺序为:气孔导度>叶温>蒸腾速率>叶片水分亏压>胞间CO2浓度,即气孔导度对净光合速率起决定性作用。
小叶楠木的多元逐步回归方程为:
该模型筛选出大气CO2浓度、蒸腾速率、胞间CO2浓度、叶片水压亏缺和空气相对湿度5个显著的影响因子,说明夏季小叶楠木的净光合速率主要受这5个因子的影响。为了进一步确定各影响因子的作用大小,进行通径分析(表7)。从直接作用绝对值看,在夏季,各因子对净光合速率的影响顺序为:叶片水分亏压>空气相对湿度>大气CO2浓度>蒸腾速率>胞间CO2浓度。即叶片水分亏压是小叶楠木净光合速率的的决定性因子。

表6 大叶楠木的净光合速率与影响因子的通径系数分析

表7 小叶楠木的净光合速率日变化与影响因子的通径系数分析
3 结论与讨论
楠木种内有大叶、小叶二种叶型,二者叶片大小、叶片厚薄、叶脉数量以及叶柄长度均存在显著差异,且同一叶型的植株上、中、下三个部位间叶片性状间也存在显著差异,尤其是叶片大小,这是植物器官的形态结构特征是与其生理功能及生长环境相适应的反映。器官结构是功能的基础,结构变化必然影响到生理生态功能的变化[22]。叶片作为暴露在环境中面积最大的器官,对环境十分敏感[23]。在长期对外界生态环境的适应中,叶片在形态结构上的变异性和可塑性是较大的[24]。楠木两种类型叶片的稳定变异造就了能适应不同环境的两种类型。
植物光合速率日变化及其影响因素一直是植物光合作用研究的重点。由多元逐步回归分析及其通径分析得到,在高温夏季,大叶楠木的净光合速率主要受气孔导度、胞间CO2浓度、叶片水分亏压、叶温和蒸腾速率等5个因子的影响,起主要作用的是气孔导度。该结果与吴林瑛等在夏季对光皮桦的研究结果基本一致[27]。而小叶楠木主要受叶片水分亏压、空气相对湿度、大气二氧化碳浓度、蒸腾速率和胞间CO2浓度等5个因子影响,且叶片水分亏压对小叶楠木的净光合速率影响最大。郎校安等发现蒸腾速率、胞间CO2浓度、气孔导度对红花深山含笑的净光合速率影响最大,而光合有效辐射、空气相对湿度、大气二氧化碳浓等环境因子对净光合速率的影响次之[26]。郭连金等研究认为两年生香果树实生苗的净光合速率与光合有效辐射和气孔导度呈显著正相关[27]。王国霞等研究认为泡桐的光合速率随天气条件、环境条件、树体发育期的不同而变化[28]。可见,因测定光合参数的环境条件、测定植株大小等不同,其对光合速率影响显著的因素也有差异。两种叶型楠木的净光合速率显著差异及其不同的影响因子与其叶片特性相关,小叶楠木叶片薄,在高温夏季易失水,叶片水分因子成为最重要的限制因素。
光合作用影响着有机物的积累量,与其生长量呈显著相关,两种叶型楠木因叶表观特征的差异,对环境的敏感性不同,通过解析影响净光合速率的主要因素,对于不同类型楠木种苗培育及其栽培具有重要作用。在高温夏季,培育楠木种苗应适当遮阴,提供充足的水分,以提高其光合速率和有机物的积累量,尤其是小叶楠木该项措施更为重要。同时,桢楠人工栽培也需在幼年期适度遮阴,可以与光皮桦等落叶树种进行混交以实现遮阴目的。
[1] 韩忠明,王云贺,林红梅,等. 吉林不同生境防风夏季光合特性[J]. 生态学报,2014,34(17):4874-4881.
[2] James B. Photosynthetic energy conversion: natural and artificial[J]. Royal Soc Chem,2009,38(1):185-196.
[3] 郑彩霞,邱箭,姜春宁,等. 胡杨多形叶气孔特征及光合特性的比较[J]. 林业科学,2006,42(8):19-24.
[4] 王海珍,韩路,徐雅丽,等. 胡杨异形叶光合作用对光强与CO2浓度的响应[J]. 植物生态学报,2014,38(10):1099-1109.
[5] 褚延广,苏晓华,黄秦军,等. 欧洲黑杨基因资源光合生理特征与生长的关系[J]. 林业科学,2010,46(7):77-83.
[6] 黄红英,窦新永,孙蓓育,等. 两种不同生态型麻疯树夏季光合特性的比较[J]. 生态学报,2009,29(6):2861-2867.
[7] 种培芳,李毅,苏世平,等. 红砂3个地理种群的光合特性及其影响因素[J]. 生态学报,2010,30(4):914-922.
[8] 卫晶晶,杨树华,郭宁,等. 不同海拔来源宽刺蔷薇的光合日变化及其生理机制[J]. 园艺学报,2013,40(8):1509-1516.
[9] 盛宝龙,常有宏,姜卫兵,等. 不同类型梨品种光合特性的比较[J]. 果树学报,2007,24(6):742-746.
[10] 张付春,伍新宇,潘明启,等. 帕米尔高原非耕地设施延晚栽培葡萄的光响应特征[J]. 果树学报,2015(4):597-603.
[11] 刘华,臧润国,张新平,等. 天山中部3种自然生境下天山雪莲的光合生理生态特性[J]. 林业科学,2009,45(3):40-48.
[12] 李俊,杨玉皎,王文丽,等. UV-B辐射增强对马铃薯叶片结构及光合参数的影响[J]. 生态学报,2017,37(16):5368-5381.
[13] 龚容,高琼. 叶片结构的水力学特性对植物生理功能影响的研究进展[J]. 植物生态学报,2015(3):300-308.
[14] Tariq A,Pan K,Olatunji O A,. Phosphorous application improves drought tolerance of.[J]. Front Plant Sci,2017,8(x):1561.
[15] Hu Y,Wang B,Hu T,Combined action of an antioxidant defence system and osmolytes on drought tolerance and post-drought recovery ofsaplings[J]. Acta Physiol Plant,2015,37(4):84.
[16] 李高志,朱亚军,周生财,等. 2种叶型桢楠对低温胁迫的生理响应及耐寒性分析[J]. 浙江农林大学学报,2017,34(2):310-318.
[17] 宗卫. CaCl2和SA对樟科3种常绿阔叶树种幼苗的抗寒性研究[D]. 荆州:长江大学,2013.
[18] Gao J,Zhang W,Li J,. Amplified fragment length polymorphism analysis of the population structure and genetic diversity of Phoebe nan (Lauraceae),a native species to China[J]. Biochem Syst Ecol,2015,64:149-155.
[19] 王茹,刘桂华,赵海燕,等. 不同光环境下桢楠幼树叶片叶绿素荧光特性[J].东北林业大学学报,2012,40(7):32-36.
[20] 胡婧楠,刘桂华,等. 2种楠木幼树光合生理特性的初步研究[J]. 安徽农业大学学报,2010,37(3):541-546.
[21] 殷国兰,谭斌,杨金亮,等. 3种珍贵用材树种1年生苗木光合特性研究[J]. 西部林业科学,2014(3):81-87.
[22] 李芳兰,包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报,2005,(S1):118-127.
[23] Wright I J,Reich P B,Westoby M. Strategy shifts in leaf physiology, structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats.[J]. Funct Ecol,2001,15(4):423-434.
[24] 朱广龙,魏学智. 酸枣叶片结构可塑性对自然梯度干旱生境的适应特征[J]. 生态学报,2016,36(19):6178-6187.
[25] 吴林瑛,邓文剑,张应中,等. 光皮桦不同无性系果期光合日变化研究[J]. 中南林业科技大学学报,2017,37(9):32-37.
[26] 郎校安,杨蕾蕾,崔铁成,等. 红花深山含笑光合速率日变化及其与环境因子的关系[J]. 西南林业大学学报,2017,37(4):22-27.
[27] 郭连金,杜佳朋,吴艳萍,等. 香果树实生苗的光合特性及其与环境因子的关系[J]. 应用生态学报,2017,28(5):1473-1481.
[28] 王国霞,关欣,刘震,等. 不同泡桐品种(种源)埋根苗生长与光合特性分析[J]. 河南农业科学,2017,46(1):116-121.
Phenotypic and Photosynthetic Properties of Leaves in
WU Xiao-lin1,YAO Yue-hua2,FANG Xia3,JIANG Zhong-long4,YANG Ping-kun1,ZHANG Mao-fu1,TONG Zai-kang3
(1. Qingyuan Experimental Forest Farm of Zhejiang, Qingyuan 323800 , China; 2. Qingyuan Forestry Bureau of Zhejiang, Qingyuan 323800 , China; 3.The State Key Laboratory of Subtropical Silviculture, Zhejiang A & F University, Hangzhou 311300, China; 4. Zhejiang Provincial Department of Forestry, Hangzhou 310020, China)
Determinations were conducted on leaf morphology and photosynthetic properties of 2-year containerf.andfseedling in 2017 in Qingyuan, Zhejiang province. It was revealed that there was significant difference between two species and even among different layers in leaf area, leaf length, leaf width, and vein number. Net photosynthetic rate off.was significantly higher than that off. Multiple stepwise regression analysis and path analysis showed that net photosynthetic rate off.was mainly influenced by stomatal conductance, intercellular CO2concentration, leaf vapor pressure deficit, leaf temperature and transpiration rate, especially stomatal conductance. While net photosynthetic rate offwas by leaf vapor pressure deficit, relative air humidity, atmospheric CO2concentration, transpiration rate and intercellular CO2concentration, especially leaf vapor pressure deficit. The experiment resulted that young trees of tested species should be shaded and have sufficient water supply in the summer.
; apparent characteristics; photosynthetic property; net photosynthetic rate
10.3969/j.issn.1001-3776.2018.03.005
S792.24
A
1001-3776(2018)03-0030-06
2018-01-09;
2018-04-15
宁波市农业重大(重点)项目(2012C10013);浙江省省院合作林业科技项(2017SY19);浙江省农业(林木)新品种选育重大科技专项(2016C02056-2)
吴小林,工程师,从事珍贵树种培育相关工作;E-mail:2925622496@qq.com。
童再康,博士,教授,从事珍贵树种遗传育种等研究工作;E-mail:zktong@zafu.edu.cn。
猜你喜欢
航空发动机(2022年4期)2022-10-13
国土绿化(2022年4期)2022-05-10
东方汽轮机(2022年1期)2022-04-13
节能技术(2022年1期)2022-03-18
西南农业学报(2021年7期)2021-08-23
江西画报(2021年6期)2021-08-19
节水灌溉(2021年5期)2021-05-26
农业技术与装备(2020年8期)2020-12-15
老友(2020年7期)2020-08-28
中国农业科技导报(2020年12期)2020-03-15
