
蒜头果中植物内生真菌烟曲霉的抗菌活性研究*
2018-08-25 07:12:50李娟陈中华华梅原晓龙虞泓王毅
西部林业科学 2018年4期
李娟,陈中华,华梅,原晓龙,虞泓,王毅
(1.云南大学中草药生物资源研究所云百草实验室,云南 昆明650091;2.国家林业局云南珍稀濒特森林植物保护和繁育重点实验室,云南省森林植物培育与开发利用重点实验室,云南 昆明650201;3.云南省林业科学院,云南 昆明650201)
植物内生真菌是一类在宿主植物的健康组织内完成整个或部分生活周期,但不引起寄主植物产生明显疾病症状的真菌[1]。几乎所有植物体内均有内生真菌的存在[2]。在健康植株中,内生真菌和其寄主植物通常是互惠共生的。植物为内生真菌提供营养和舒适的环境,有些内生真菌能促进寄主植物的生长并提高植物对生物逆境和非生物逆境的耐受性[3-4]。草本植物中的内生真菌能够产生具有毒性的生物碱,从而保护寄主不被食草动物取食[5]。香柱菌属 (Epichloe)一内生真菌新种 (Epichloe gansuensis),可以抑制醉马草 (Achnatherum inebrians)上禾本科布氏白粉菌 (Blumeria graminis)的定殖[6]。牧草内生菌产生的毒麦碱对邻近植物有较强的化感作用。面对非生物胁迫,植物内生真菌能产生相应的代谢产物,提高寄主对干旱、高温、高盐、重金属等胁迫的耐受性。由于植物内生真菌生存环境的多样性以及种类的多样性,决定了植物内生真菌代谢产物的多样性。研究表明,从植物中分离得到的内生真菌可产生多种活性物质,多具有抗肿瘤、抗菌、抗病毒、杀虫、免疫抑制、抗氧化和降糖等活性,其中有一些是具有特殊活性的新物质[7]。如短叶紫杉 (Taxus brevifolia)的树皮中分离出产紫杉醇的内生真菌 (Taxomyces andreanae)能产生抗乳腺癌、子宫癌、卵巢癌的化合物紫杉醇 (taxol)[8]。从蛇足石杉 (Huperzia serrata)的内生菌 (Penicillium chrysogenum P1X)中分离得到9种具有较罕见C25类固醇骨架的抗菌甾体类化合物[9]。
目前临床上抗生素的大量使用,使许多致病细菌的耐药性不断增强,根据2012年中国CHINET细菌耐药性检测报道,耐甲氧西林凝固酶阴性葡萄球菌 (MRCNS)的检出率平均为77.1%;大肠埃希菌 (Escherichia coli)对环丙沙星、庆大霉素和哌拉西林的耐药率均接近或高于50.0%[10]。耐药菌感染已严重危害人类健康,寻找新的抗菌药物来代替抗生素已迫在眉睫。植物内生真菌产生的抗菌物质多数是新化合物,因此,有望从中寻找到新的抗生素。例如,从红树林植物卤蕨 (Acrostichum aureurm)中分离得到的内生真菌 (Penicillium sp.0935030)能产生两种抑制耐甲氧西林金黄色葡萄球菌 (MRSA)的环肽化合物[11]。从鸡冠刺桐(Erythrina crista-galli)的内生真菌 (Phomopsis sp.)发酵液中获得的一个新抗生素Phomol对24种细菌和真菌都显示出很好的抗菌活性[12]。
目前,虽然已从其他植物中分离获得具有许多生物活性的内生真菌,但对云南珍稀濒危植物蒜头果 (Malania oleifera)内生真菌的活性研究还未见报道。单菌多产物 (OMSAC)策略主要是通过改变培养条件、培养方法等手段,激活微生物中沉默基因,以产生大量具有活性的代谢产物,为新型药物的研发提供先导化合物[13]。因此,本研究从我国特有的单属种植物,铁青树科 (Olacaceae)蒜头果属 (Malania)的蒜头果中分离获得植物内生真菌,使用OMSAC策略进行发酵,并检测其发酵产物的抗菌活性,以期为蒜头果内生真菌的统合利用提供参考。
1 材料与方法
1.1 试验材料
烟曲霉菌种 (Aspergillus fumigatus)从蒜头果植株中分离获得,经ITS及形态鉴定后,保存在云南省林业科学院,国家林业局云南珍稀濒特森林植物繁育和保护重点实验室,菌株编号为MOEF013。供试6种人体致病细菌分别为蜡样芽孢杆菌 (Bacillus cereus)、副溶血性弧菌 (Vibrio parahaemolyticus)、无乳链球菌 (Streptcococcus agalactiae)、福氏志贺氏菌 (Shigella flexneri)、金黄色葡萄球菌(Staphylococcus aureus)、藤黄微球菌 (Micrococcus luteus)。由昆明市食品药品检验所赠送。
1.2 不同液体培养基的配制
OMAM培养基 蔗糖 (20g/L)与葡萄糖(20g/L)为碳源,酵母粉 (1g/L)和大豆蛋白胨(20g/L)为氮源。
不同氮源的培养基 麦芽糖 (1.8g/L)与葡萄糖 (6g/L)为碳源,编号为M与G,分别以4g/L的麦芽浸粉 (M1)、酵母粉 (Y2)、番茄浸粉(T)、马铃薯浸粉 (P)、大豆蛋白胨 (S)、胰蛋白胨 (Y1)、牛肉浸粉 (B)、酪蛋白胨 (L)和香蕉粉 (B1)为氮源,配制成碳源相同氮源不同的液体培养基,编号分别为 MGM1、MGY2、MGT、MGP、 MGS、 MGY1、 MGB、 MGL、 MGB1。
不同碳源的培养基 麦芽浸粉 (6g/L)与酵母粉 (3g/L)为氮源,分别以4g/L的甘露醇、麦芽糖、山梨醇、蔗糖、乳糖、葡萄糖、肌醇、果糖、可溶性淀粉为碳源,配制成氮源相同碳源不同的液体培养基,编号分别为man、mal、sor、suc、luc、 glu、 ino、 fru、 sta。
1.3 OSMAC策略寻找最佳培养条件
不同条件下分别培养MOEF013,对其不同的提取物做活性检测,具体为无菌条件下,研碎烟曲霉菌体;不同接种量为研碎15mg、30mg、60mg、90mg、120mg菌体各一份,在OMAM液体培养基中培养;不同培养条件为接种到OMAM培养基摇床培养5d后,分别加入细菌共培养、周期性改变培养温度 (15℃与28℃交替改变,每12h改变一次)、共培养的同时改变培养温度3种条件下再培养5d;不同培养基分别接种到不同碳源和不同氮源的液体培养基中,28℃,150r/min摇床培养10d。
收获培养物完全干燥后加入二甲基亚砜(Dimethyl sulfoxide,DMSO)制备成50mg/mL提取物的DMSO溶液,备用。
1.4 提取物抗菌活性检测
致病细菌悬浮液的制备 分别接种6种检测细菌于LB固体培养基中,37℃恒温培养12h后,挑取单菌落接种到5mL液体LB培养基中,摇床37℃,180r/min培养至菌液浓度达到1×106-6×106cfu/mL,备用。
抑菌实验采用纸片法 将各检测菌液均匀涂布在LB固体培养基上,略干后将加有10μL提取物的滤纸片 (φ=5mm)放置在含有检测菌的培养基上,10μL DMSO作为阴性对照,37℃恒温培养24h后测量抑菌圈大小。每个菌种重复3次实验。
测定最小抑菌活性 (MIC) 采用二倍稀释法,测定具有活性提取物的MIC值。
2 结果与分析
2.1 最低抑菌浓度
检测MOEF013提取物对6种人体致病菌的抗菌活性,结果显示,提取物对蜡样芽孢杆菌、副溶血性弧菌、无乳链球菌、福氏志贺氏菌、金黄色葡萄球菌、藤黄微球菌具有良好的抗菌活性。二倍稀释法测定MOEF013发酵提取物对6种致病菌的MIC(表1),提取物对副溶血性弧菌和藤黄微球菌抑菌效果较好,最低抑菌浓度为6.25mg/mL,金黄色葡萄球菌、无乳链球菌、福氏志贺氏菌最低抑菌浓度为12.5mg/mL。蜡样芽孢杆菌的MIC值为25.0mg/mL,说明蜡样芽孢杆菌对烟曲霉提取物敏感性较低。

表1 MOEF013提取物对6种致病菌的MICTab.1 The minimum inhibitory concentration of MOEF013 extracts to 6 kinds of pathogenic bacteria
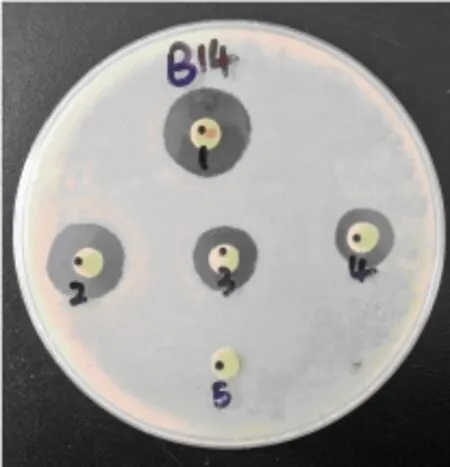
图1 MOEF013提取物对藤黄微球菌的MICFig.1 MIC of MOEF013 extracts to Micrococcus luteus
2.2 接种量对烟曲霉抑菌活性的影响
按照不同接种量培养MOEF013菌株10d后收获其代谢产物,乙酸乙酯提取后测定抑菌活性。结果显示,不同接种量对抑菌活性没有影响,但对产物的提取量有明显影响 (图2)。

图2 接种量对MOEF013次生代谢产物提取量的影响Fig.2 Influence of inoculation quantity on the extraction of secondary metabolites from MOEF013
由图2可以看出,随着接种量的增加,提取量逐渐增加,当接种量为60mg时,提取量最高,接种量大于60mg时,提取量逐渐下降。
2.3 共培养和培养温度变化对MOEF013次生代谢产物抑菌活性的影响
通过周期性改变培养温度、与细菌共培养后发现烟曲霉提取物抑菌活性有所变化 (表2)。相比对照组,温度变化后烟曲霉提取物除对无乳链球菌抗菌效果不变,对藤黄微球菌抗菌效果略微下降外,对其余4种致病菌的抗菌活性都有所增强。加入细菌共培养后对蜡样芽孢杆菌、金黄色葡萄球菌、藤黄微球菌的抗菌活性有所增强,但对副溶血性弧菌、无乳链球菌、福氏志贺氏菌的抗菌活性下降。共培养并改变温度后对蜡样芽孢杆菌、副溶血性弧菌、福氏志贺氏菌3种致病菌的抗菌效果有所增强,但对无乳链球菌、藤黄微球菌的抗菌效果减弱,对金黄色葡萄球菌没有影响。
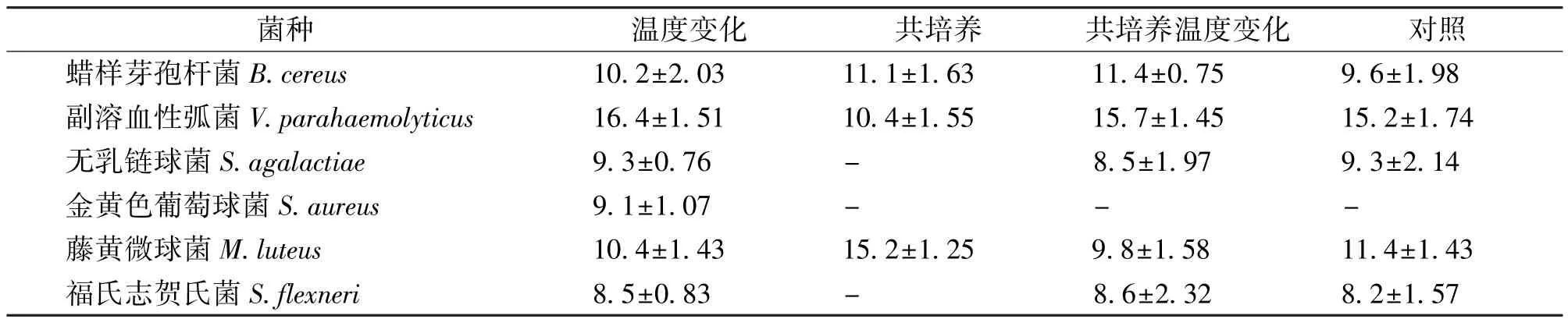
表2 共培养和温度变化对MOEF013次生代谢产物抑菌活性影响Tab.2 Influence of co-culture and temperature changes on the antibacterial activity of secondary metabolites from MOEF013
2.4 不同培养基对MOEF013次生代谢产物抑菌效果的影响
2.4.1 不同氮源对MOEF013提取物抑菌效果的比较
MOEF013在9种不同氮源培养基中培养后经乙酸乙酯萃取所得提取物对6种致病细菌的抑菌效果差别较明显 (图3)。可以看出不同培养基对MOEF013的次生代谢产物有影响。其中MGY1和MGP培养基培养MOEF013产生的次生代谢产物对6种致病菌抗菌活性均较好。但MGP培养基培养下提取物的收获量 (31.2mg)比MGY1培养基的收获量 (17.1mg)更高,所以确定马铃薯粉为培养蒜头果内生真菌烟曲霉的最佳氮源。此外,不同氮源下提取物对不同致病菌的抗菌效果也有差异。除提取物MGM1对金黄色葡萄杆菌没有抗菌活性,MGS提取物对金黄色葡萄杆菌和福氏志贺氏菌没有抗菌活性外,其余氮源所获得提取物对6种致病细菌均有抗菌活性。MGL提取物对金黄色葡萄球菌抗菌效果最好,抑菌圈直径达19mm。MGB1提取物对藤黄微球菌抗菌效果较其他氮源都好。副溶血性弧菌对MGY1,MGM1,MGB1提取物较敏感(图4)。

图3 不同氮源培养基下MOEF013次生代谢产物抑菌活性比较Fig.3 Antibacterial activity of MOEF013 secondary metabolites in different nitrogen sources
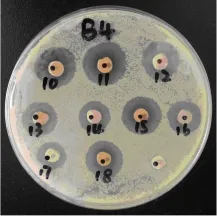
图4 不同氮源培养基发酵提取物对副溶血性弧菌抑菌效果Fig.4 Antibacterial activity of MOEF013 extracts in different nitrogen sources to Vibrio parahaemolyticus
2.4.2 不同碳源对MOEF013提取物抑菌效果的比较
分别在9种不同碳源培养基中培养MOEF013后所得提取物对6种致病细菌的抑菌效果有差异(图5)。

图5 不同碳源培养基下MOEF013次生代谢产物抑菌活性比较Fig.5 Antibacterial activity of MOEF013 secondary metabolites in different carbon sources
glu为碳源的提取物对6种致病菌均有抑菌活性。mal为碳源的提取物对5种致病细菌有抑菌活性,尤其对副溶血性弧菌抑制效果显著,但对福氏志贺氏菌没有抑菌活性。suc培养基所获提取物对除藤黄微球菌外的5种致病菌均有抗菌效果,对无乳链球菌抗菌效果较明显。man和sor提取物对4种致病菌有抗菌活性,对藤黄微球菌和福氏志贺氏菌没有活性。luc为碳源所获提取物对蜡样芽胞杆菌和福氏志贺氏菌没有抗菌活性,对其余致病菌均有抑制效果。fru提取物对藤黄微球菌和福氏志贺氏菌没有抑制效果。sta为碳源的提取物对蜡样芽胞杆菌和无乳链球菌没有抑菌作用。ino为碳源所获得提取物抗菌活性较差,仅对蜡样芽胞杆菌和副溶血性弧菌有抑制作用。
3 结论与讨论
蒜头果为我国特有植物,被列为国家二级重点保护植物,仅分布于我国云南东南部至广西西部的狭窄地带,不仅分布极其狭窄,且现存资源量十分稀少[14]。本研究从蒜头果树皮中成功分离获得植物内生真菌—烟曲霉,对其内生真菌烟曲霉发酵后提取物的抗菌活性进行检测,发现它对6种人体致病细菌 (蜡样芽孢杆菌、副溶血性弧菌、无乳链球菌、福氏志贺氏菌、金黄色葡萄球菌、藤黄微球菌)均有抑菌活性,其中对副溶血性弧菌、藤黄微球菌最低抑制浓度达到6.25mg/mL。植物内生真菌与宿主植物的共生关系是经历了漫长的进化过程后相互适应的结果,其代谢产物的种类与其宿主植物也息息相关[15]。不同属的植物因其内环境的不同,故寄生于不同属植物的同属内生真菌代谢产物也会有所不同。例如从禾本科 (Gramineae)植物狗牙根 (Cynodon dactylon)中获得的内生真菌(Aspergillus sp.)能产生3种具抗菌活性的甾类化合物,而从红豆杉 (Taxus mairei)植物中获得的内生真菌 (Aspergillus clavatonanicus)能产生对多种植物病原菌都有抑制作用的内酯化合物 patulin[16]。此外,植物内生真菌次生代谢产物也受到发酵条件的影响。林淑婷等发现,不同发酵条件下,海南粗榧 (Cephataxus hainanensis Li.)中一种内生真菌的代谢产物种类和产量均有明显差异[17]。因此,本报道通过OSMAC策略,探究了我国濒危特有物种蒜头果内生真菌MOEF013菌株的最佳发酵条件,寻找到使其代谢产物活性最好、产量最高的培养条件,在该条件下,MOEF013菌株次生代谢产物对蜡样芽孢杆菌、副溶血性弧菌、无乳链球菌、福氏志贺氏菌、金黄色葡萄球菌、藤黄微球菌均有良好的抑菌作用。在以往的文献中,仅见到烟曲霉次生代谢产物对金黄色葡萄球菌、白假丝酵母菌 (Candida albicans)抑菌活性的报道[18-20]。由新的抗菌活性可以推测,蒜头果内生真菌MOEF013菌株在一定条件下可能产生了新的抗菌化合物。
此外,本研究发现MGM1培养基发酵下所得提取物对副溶血性弧菌抑菌效果较好,但对金黄色葡萄球菌没有抑菌效果,而MGL所得提取物对金黄色葡萄球菌抑菌效果明显优于其他致病菌,因此可以推断,蒜头果内生真菌烟曲霉所产生的抗菌成分不止一种。类似的情况在其他微生物中也存在,如谢海平等发现海洋枯草芽孢杆菌 (Bacillus subtilis)Bs-1在PDA培养基中发酵至少产生3种抗真菌活性化合物[21]。绿僵菌 (Metarhizium anisopliae)除了能产生具有抗菌效果的烟曲霉酸 (helvolic acid),还能产生一种烟曲霉酸的衍生物 (1,2-dihydrohelvolic acid)[22]。但本研究中所用的样品仅为蒜头果内生真菌烟曲霉的发酵液粗提物,对于起到抗菌作用的究竟有几种化合物、分别是什么化合物还有待进一步的鉴定和研究。在今后的研究中,将大量发酵蒜头果内生真菌烟曲霉,对其提取物进行分离、纯化、鉴定等进一步研究,确定提取物中抗菌活性成分,并深入研究其抗菌的机制,以期为新型抗菌药物提供先导化合物。
猜你喜欢
湖北农机化(2020年4期)2020-07-24 09:07:16
临床医药文献杂志(电子版)(2020年23期)2020-02-28 03:59:52
世界农药(2019年4期)2019-12-30 06:25:10
今日农业(2019年11期)2019-08-15 00:56:32
乡村地理(2018年2期)2018-09-19 06:44:02
中国酿造(2016年12期)2016-03-01 03:08:11
吉林大学学报(医学版)(2015年3期)2015-12-17 07:47:42
安徽中医药大学学报(2014年2期)2014-06-19 13:22:16
长江大学学报(自科版)(2014年23期)2014-03-27 10:51:57
中国酿造(2014年9期)2014-03-11 20:21:03
