
生物结皮对高寒退化草地植物群落的影响
2018-08-23 11:52徐恒康
草地学报 2018年3期
徐恒康, , , , , , *
(1. 中国农业大学动物科技学院, 北京 100193; 2. 青海贵南县草原站, 青海 贵南 730070)
生物结皮是藻类、地衣、苔藓真菌和细菌与土壤颗粒通过团聚和胶结作用在土壤表层形成的复合体[1]。生物土壤结皮具有防风固沙,防止水土流失,减慢水分蒸发的生态功能;能够固定大气中的碳、氮,增加土壤有机质,对微管植物微量元素的富集和营养物质的循环起到重要作用[2-4]。地衣和苔藓结皮是生物结皮的重要组成部分,通常出现在生物结皮发育成熟阶段[5]。近几十年,国内外学者对地衣和苔藓结皮的形态结构、分布格局、生物组成、特征分类和生态功能和管理等方面开展了研究[6]。生物结皮与微管植物间存在怎样的相互关系也是当前研究的热点,在不同的环境下,生物土壤结皮可以促进和抑制种子萌发和植物定植[7]。生物土壤结皮与当地本土植物关系紧密且可防止外来物种的入侵,当生物结皮大量生长时,结皮的生态结构和物种组成发生改变并抑制本土物种的生长[8-9]。一些研究认为,生物结皮促进了植物的生长,并提供植物所需要的营养物质[10];也有学者认为苔藓植物迅速吸收水分和养分,与维管植物竞争资源[11];此外,也有观点认为两者占据不同的生态位,没有明显的互惠和竞争的关系[12]。生物结皮对植物群落的群落组成变化、各类型群落的特征对比以及生物结皮盖度与群落盖度的关系尚不明确。尽管对生物结皮与维管植物关系的报道很多,但多集中于荒漠地区和沙漠地区的研究[5, 13-14],对于青藏高原高寒区域,生物结皮与植物群落的关系是促进或是抑制作用尚不明晰。本文探究青藏高原高寒退化草地生物结皮对植物群落结构以及多样性的影响,分析了生物结皮盖度与草本植物盖度之间的相关性分析,以期为退化高寒草地潜在的植物演替方向与生态恢复机制提供科学依据。
1 材料与方法
1.1 研究区概括
研究区域位于青海省海南藏族自治州贵南县草原站(100°13′~101°33′ E,35°09′~36°08′ N),属高原大陆性气候,年平均气温为2.3℃,年降水量为403.8 mm,年平均蒸发量为1 378.5 mm。植被类型为针茅化高寒草原,主要优势种有矮生嵩草(Kobresiahumilis)、垂穗披碱草(Elymusnutans)、溚草(KoeleriacristataPers)和紫花针茅(StipacapillataLinn)等。土壤类型以高山草甸土为主,在草本植物之间镶嵌着发育良好的生物结皮,其中苔藓结皮占优势,偶见地衣结皮。

图1 左一为苔藓结皮,右一为地衣结皮Fig.1 Moss biocrust in left and lichen biocrust in right
1.2 群落调查与取样方法
于2017年7月-9月开展野外取样调查,试验地位于贵南县森多乡扎什给村,试验地围栏200 m×300 m,在围栏样地内按对角线设置3 m×3 m的样方30个。在每个样方随机选取1个50 cm×50 cm的样方,共30个。用铁丝方格网(由256个面积为 1.25 mm × 1.25 mm 小格构成)调查结皮盖度。在每个调查样方中,记录不同结皮出现的格子总数,并根据不同结皮所占铁丝网的格数推算出盖度。调查样方群落内草本植物的种类、高度、盖度、株数,其中草本植物群落鉴定到种。按照苔藓、地衣的形态特征分别记录样方内苔藓地衣的存在情况,统计整理后分为类型Ⅰ(地衣结皮和苔藓结皮都存在)、类型Ⅱ(只有苔藓结皮存在)、类型Ⅲ(苔藓和地衣结皮均无)三种样方类型,其中地衣结皮和苔藓结皮同时存在的样方(类型Ⅰ)13个;只有苔藓结皮存在的样方(类型Ⅱ)13个,样方内地衣和苔藓结皮都无的样方(类型Ⅲ)4个(其中结皮盖度达到2%以上才算出现,只有地衣存在的样方由于地衣盖度很小,因此忽略不计)。
1.3 重要值和多样性指数
重要值计算公式:重要值=(相对高度+相对盖度+相对频度)/3
采用丰富度指数和Shannon-Wiener指数进行多样性测定,计算公式如下:
(1)Margalet丰富度指数:R= (S-1)/lnN
(2)Shannon-Wiener指数
H=∑(-PilnPi)
上述各式中,Pi为物种i的重要值,S为物种i所在样方的物种总数,N为样方内所有植物总数。
1.4 数据分析
利用Excel 2003对植物种类进行统计和分析,对不同处理间的植物种数、盖度、平均高度、物种丰富度、Shannon-Wiener指数等进行差异性分析,探究生物结皮对高寒草地植物群落的影响。
利用SPSS16.0对结皮盖度和植物群落盖度做相关性分析,利用曲线估计(curve estimation)进行拟合,找出有意义且拟合度最优的曲线和方程(决定系数采用校正后R2值,R2最大, 且P<0.05),利用Sigmaplot 12.0回归分析并作图,探究不同生物结皮盖度与植物群落盖度之间的关系。
2 结果与分析
2.1 植物群落物种组成
试验区内草本植物共23种,其中2种为短命植物,分属于9个科(表2)。苔藓和地衣结皮都存在的生境下(Ⅰ)植物种类共计7科20种植物,仅有苔藓结皮存在的样方生境下(Ⅱ)植物种类共计8科21种。苔藓地衣结皮都不存在的生境下(Ⅲ)的植被种类共计7科16种。结皮存在样方内植被种数比对照样方多1科4种,其中Ⅰ类型中豆科、禾本科和莎草科所占比例50%,杂草比例为50%;Ⅱ类型中豆科、禾本科和莎草科所占比例47.61%,杂草比例为52.39%。Ⅲ类中没有豆科出现,禾本科和莎草科所占比例为43.75%,杂草比例为56.25%。其中玄参科只在Ⅰ,Ⅲ中出现,唇形科只在Ⅱ,Ⅲ中出现(见图2)。
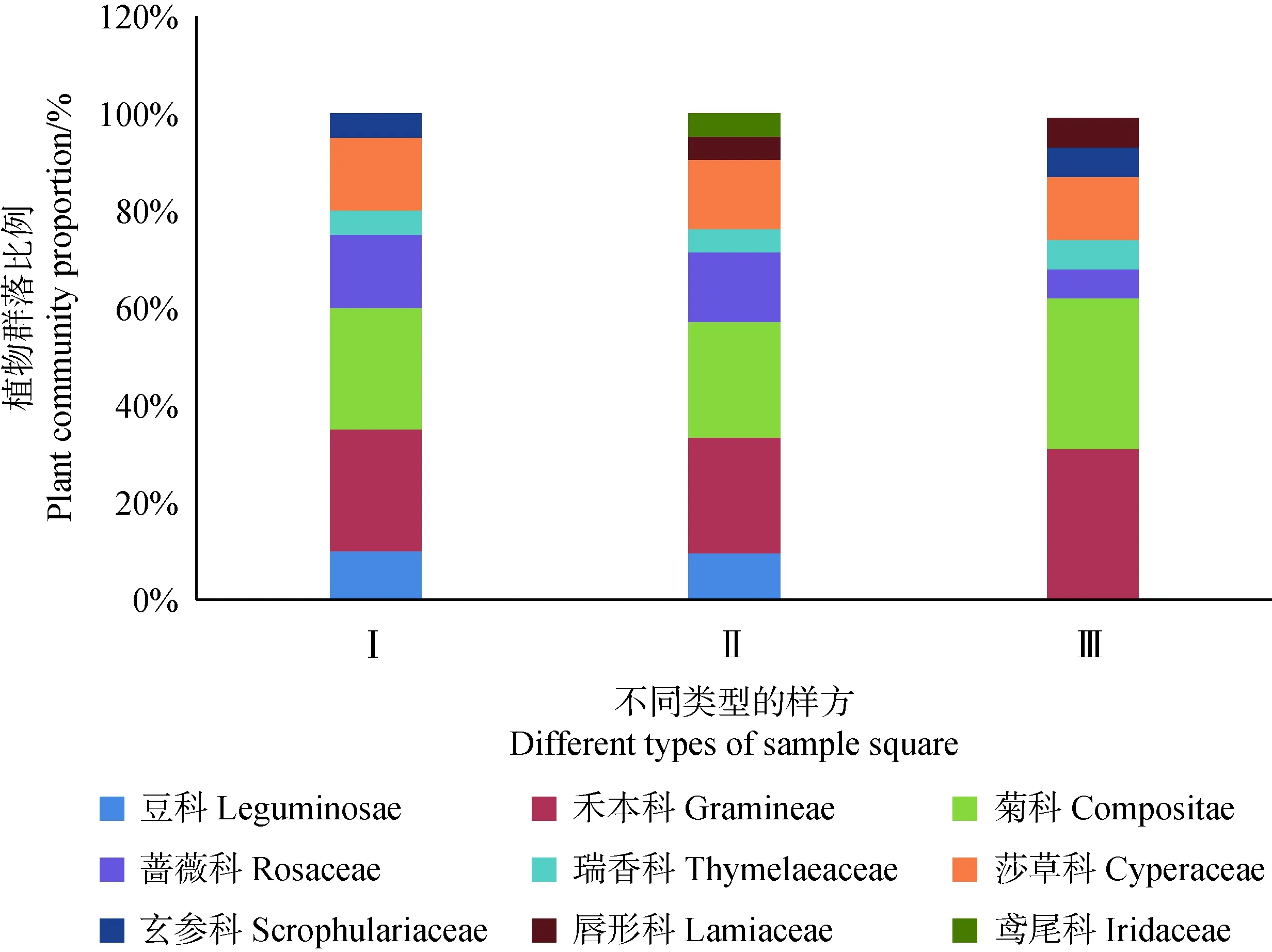
图2 结皮样方内植物群落组成Fig.2 Community structure of herbaceous plants around different types of crusts
2.2 生物结皮对植物群落盖度和多样性的影响
生物结皮显著影响了植物群落的盖度、高度以及多样性(表1),类型Ⅰ样方的(地衣和苔藓结皮都存在)草本植物的盖度和植被平均高度都显著高于类型Ⅱ(仅有苔藓结皮存在)(P<0.01),但是类型Ⅰ、类型Ⅱ与类型Ⅲ(没有结皮存在)样方中植被盖度与植被平均高度差异均不显著;Ⅱ类型样方中的苔藓结皮盖度显著高于Ⅰ类型样方(F=11.33,P<0.01);类型Ⅲ(没有结皮存在)的样方其丰富度指数和Shannon-Wiener指数显著高于类型Ⅰ(苔藓和地衣都存在)(P<0.05),而与Ⅱ类型差异不显著。

表1 不同类型生物结皮对植物群落多样性的影响Table 1 The effects of different types of biological crusts on plant community diversity
注:不同小写字母表示有显著性差异(P<0.05)
Note:Different lowercases indicate the significance at the 0.05 level
2.3 群落重要值变化
由表2可以看出生物结皮对群落物种组成产生了影响,植物物种重要值有所改变。研究发现(表2),溚草(KoeleriacristataPers)、草地早熟禾(PoapratensisL)、紫花针茅(Stipapurpurea)与矮火绒(Leontopodiumnanum)为试验样地的优势种。溚草重要值在三个类型的样方中比较;类型Ⅱ>类型Ⅰ>类型Ⅲ;草地早熟禾的变化趋势为:类型Ⅱ>类型Ⅲ>类型Ⅰ;紫花针茅的重要值在类型Ⅰ中最高(0.114);矮火绒重要值变化趋势与溚草一致。

表2 不同类型生物结皮中植物物种的重要值Table 2 Important value of plant species in different types of biological crusts
注:*表示短命植物
Note:*indicates the shortlife plant
2.4 生物结皮盖度与群落盖度的关系
植物群落盖度与苔藓结皮盖度之间也存在着相关关系。分析得出:在类型Ⅰ中,苔藓结皮的盖度与草本植物盖度为正相关关系(F=4.084,R2=0.2707,P=0.048;图3a),地衣结皮盖度与草本植物盖度无显著相关性(F=0.906,R2=0.076,P=0.076);在类型Ⅱ中(只有苔藓结皮存在),苔藓结皮盖度与草本植物盖度呈现极显著正相关(F=10.087,R2=0.4783,P=0.009;图3b)。由此可见,苔藓生物结皮的盖度与草本植物盖度在有地衣和地衣共存的样方中显著正相关(P<0.05),在仅有苔藓结皮存在的样方中极显著正相关(P<0.01)。
3 讨论
生物结皮可通过侵蚀和沉积以改变水分分配和养分分布格局,生物结皮的分解可以增加土壤的有机质含量,这些都会影响植物的生长发育以及土壤-植物生态系统的演变[15-16]。生物结皮与草地植物群落有着直接或间接的相互关系,大多数学者[17-18]认为结皮盖度与植物群落盖度之间存在正相关关系,而有些学者[19-20]认为它们存在负相关关系,即生物结皮的存在不利于荒漠植物的萌发和定居,此外有观点认为两者占据着不同的生态位,没有明显的互惠或竞争的关系[12]。生物结皮对种子植物多样性的影响,是由于生物结皮的存在对于土壤种子库、种子萌发以及植物存活等综合影响的结果[21]。
本研究发现,生物结皮对草本植物的群落结构产生一定程度的影响。其中Ⅰ类型中豆科、禾本科和莎草科所占比例50%,杂草比例为50%;Ⅱ类型中豆科、禾本科和莎草科所占比例47.61%,杂草比例为52.39%。Ⅲ类型中没有豆科出现,禾本科和莎草科所占比例为43.75%,杂草比例为56.25%。没有结皮存在的样方的物种丰富度指数和Shannon-Wiener指数显著高于苔藓和地衣都存在的类型Ⅰ。可以得出:生物结皮的存在降低了杂草的比例,且初级发育阶段的生物结皮(地衣和苔藓共存)显著减少了植物群落的多样性,而高级发育阶段的生物结皮(仅有苔藓结皮存在)对植物群落的多样性降低程度不显著。这与前人[22-24]在荒漠和沙漠研究结果一致。庄伟伟等[22, 25]研究发现,对生物结皮层进行小范围,适宜的破坏,将会提高维管植物定居的可能性,从而说明发育良好的生物结皮存在着抑制维管植物生长的可能性。可能是因为生物结皮对种子的“隔离”作用,由于发育良好的生物结皮在土壤表层形成坚实的“外壳”,导致生物结皮的种子不能定植,从而造成对植物群落结构和物种多样性的差异。研究还发现,溚草、草地早熟禾、紫花针茅与矮火绒为实验样地的优势种,然而在没有结皮存在的样方中垂穗披碱草作为优势种出现。可以初步推测生物结皮的存在抑制了高寒草地中垂穗披碱草的定植与生长。可能由于披碱草的繁殖方式是种子生殖,而且披碱草的种子体积较大,且对比发现溚草、草地草熟禾、紫花针茅与火绒草都是密丛型或者垫装丛生型,占据了一定的绝对空间优势,而披碱草则为疏丛型多年生植物。因此,生物结皮影响了高寒草地多年生疏丛型草本植物的分布与生长。分析生物结皮盖度与草本植物盖度的相关性联系,得出苔藓生物结皮与草本植物盖度呈正相关关系。可能是因为生物结皮积累了大量植物生长所需的必要元素,促进了植物吸收和成长,而草本植物给苔藓结皮的生长提供了遮阴和减少蒸发的微环境,从而促进苔藓结皮的生长[10]。可能其中存在着一个“阈值”达到一定程度后苔藓植物与维管植物竞争资源,植被与苔藓结皮的盖度呈负相关[26]。苔藓结皮和地衣结皮有着不同的微生物群落,也可能是因为微生物的多样性的差异直接或间接影响植物群落[27]。然而,生物结皮可能在其前期发育阶段(苔藓和地衣都存在)显著降低了植物群落多样性和丰富度指数,而在其高级发育阶段,对于植物群落多样性和丰富度指数降低作用影响不显著[28]。原因可能是生物结皮和植物群落在演替过程中达到了相对的平衡的稳定状态。地衣结皮的盖度与微管植物关系不明显,其原因可能因为地衣不需要进行光合作用,需要的资源也较少,与植物的盖度关系不明显。

图3 草本植物盖度与苔藓植物盖度的相关分析Fig.3 Linear relationships between biological soil crust coverage and vascular plant coverage
4 结论
在青藏高原高寒退化草地生态系统中,生物结皮的存在影响了植物的群落结构。生物结皮的存在降低了杂草的比例,且初级发育阶段的生物结皮(地衣和苔藓共存)显著降低了植物群落的多样性,而高级发育阶段的生物结皮(仅有苔藓结皮存在)对植物群落的多样性降低程度不显著;苔藓生物结皮的盖度与草本植物盖度呈正相关关系,在地衣和苔藓结皮共存的样方中正相关显著(P<0.05),在仅有苔藓结皮存在的样方中正相关关系极显著(P<0.01)。生物结皮在高寒草地生态系统中扮演着重要的角色,可能在调节高寒退化植物群落的结构和功能方面发挥重要作用。
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04
林业勘查设计(2022年1期)2022-02-15
土壤(2021年5期)2021-11-17
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
黑龙江工程学院学报(2020年5期)2020-10-21
林业调查规划(2020年3期)2020-06-03
水土保持通报(2020年6期)2020-03-10
水土保持研究(2019年6期)2019-10-19
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10
