
二氯化钴诱导缺氧对肌细胞萎缩的调控机制研究
2018-08-13 05:43陈睿佘燕玲江婷周珊瑶史华彩黎程
中国运动医学杂志 2018年4期
陈睿 佘燕玲 江婷 周珊瑶 史华彩 黎程
1广东省第二人民医院,广东省传统医学与运动伤害康复研究所(广东广州 510317)2中山大学附属第三医院放射科(广东广州 510630)
氧是细胞生存及能量代谢的重要底物,缺氧可影响机体代谢功能及细胞存活。缺氧诱导的肌萎缩可见于过度训练、高原低氧、下肢动脉闭塞症和慢性非阻塞性肺疾病等[1-3]。近些年来研究认为,自噬参与骨骼肌细胞内稳态平衡及肌萎缩相关疾病的生理病理过程[4,5]。自噬是细胞通过形成双层膜结构的自噬体,将自身受损的细胞器和大分子物质包裹并运送至溶酶体进行分解代谢的过程。适当的自噬有利于细胞在不良环境中进行防御,而过量的自噬则可能导致细胞死亡[6]。本实验探讨二氯化钴(cobaltous chloride,CoCl2)诱导缺氧对肌细胞萎缩的调控机制。
1 材料与方法
1.1 主要仪器与材料
CO2培养箱:赛默飞世尔科技公司。多功能酶标仪:Bioteck公司。透射电子显微镜:日本电子株式会社。荧光定量PCR仪:美国ABI公司。电泳转印系统:北京凯元公司。化学发光成像系统:上海天能科技有限公司。高糖DMEM培养基购于Gibco。胎牛血清、马血清购于Hyclone。CoCl2购于Sigma。3-Methyladenine(3MA)购于Selleck。吉姆萨染色液、ECL发光试剂盒购于南京凯基生物科技发展有限公司。活性氧(reactive oxygen species,ROS)检测试剂盒、诱导因子-1 α(hypoxia-inducible factor-1α,HIF-1α)、Bcl2/腺病毒E1B 19kDa相关蛋白3(Bcl2/adenovirus E1B19kDa interacting protein 3,BNIP3)、肌肉萎缩盒F基因(muscle atrophy F-box,MAFbx)一抗购于abcam。微管相关蛋白1轻链-3(microtubule associated protein1 light chain 3,LC3)、tubulin一抗、辣根过氧化物酶标记山羊抗兔购于ABclonal。转录试剂盒及聚合酶链反应(PCR)扩增试剂盒购自Takara公司。引物由上海生工生物工程有限公司合成。
1.2 细胞培养
C2C12小鼠骨骼肌细胞系购买于中国科学院干细胞库,使用含10%胎牛血清的高糖DMEM培养基,置于37℃,体积分数5%CO2的培养箱中进行培养,隔天换液,观察细胞形态和生长情况。当细胞达到90%~100%融合时吸去完全培养液,使用含2%马血清的高糖DMEM进行诱导分化,隔天换液。5天后观察细胞分化为多核长条状肌管。正常组不加药物处理,CoCl2组加入200 μM CoCl2诱导缺氧,正常+3MA组加入5 mM 3MA,CoCl2+3MA组加入200 μM CoCl2及5 mM 3MA。24小时后收集细胞,进行后续实验。
1.3 多功能酶标仪检测ROS含量
将细胞培养于96孔黑色底透培养板中,诱导细胞缺氧后去除培养液。加入100 μl 1×Buffer清洗细胞。去除Buffer,加入100 μl 浓度为20 μM DCFDA溶液,于37℃避光孵育45分钟后移除DCFDA溶液,加入100 μl 1×Buffer,使用多功能酶标仪于488 nm激发波长,525 nm发射波长进行检测。
1.4 吉姆萨染色
使用预冷甲醇固定细胞10分钟,磷酸缓冲盐溶液(phosphate buffer saline,PBS)清洗后使用吉姆萨染色液染色20分钟,自来水冲洗。晾干后置于光学显微镜下观察拍照。
1.5 透射电镜观察自噬体形成情况
收集细胞后,2.5%戊二醛中前固定4小时,经PBS清洗后放入1%锇酸中后固定2 h,梯度乙醇(50%、70%、80%、90%、100%)依次脱水,epson812中进行包埋。包埋后放入烤箱中聚合72小时,之后于超薄切片机上进行切片,再经硝酸铅和醋酸铀双重染色,使用JEM-1200EX型透射电镜观察并拍照。
1.6 实时荧光定量PCRPCR检测相关基因的表达
加入1 ml TRIzol于6孔板中裂解细胞,室温放置5分钟后提取RNA并使用NanoDrop2000进行RNA浓度及质量测定。反转录按照Takara试剂盒说明书进行。10 μl反应体系如下:5×PrimeScriptRT Master Mix 2 μl,total RNA 1 μl(500 ng),DEPC 水补足到10 μl。PCR条件:95℃ 30 s,95℃ 5 s,60℃ 30 s,40个循环。引物序列如下:18 s上游:GTAACCCGTTGAACCCCATT;18s下 游 :CCATCCAATCGGTAGTAGCG;MAFbx上游:GAGTGGCATCGCCCAAAAGA;MAFbx下游:TCTGGAGAAGTTCCCGTATAAGT;HIF-1α上游:ACCTTCATCGGAAACTCCAAAG;HIF-1α下游:CTGTTAGGCTGGGAAAAGTTAGG;BNIP3上游:TGAATCTGGACGAAGTAGCTCC;BNIP3下游:CAGACGCCTTCCAATGTAGATC;LC3上 游 :CGATACAAGGGGGAGAAGCA;LC3下游:ACTTCGGAGATGGGAGTGGA。
1.7 免疫蛋白印迹检测相关蛋白的表达
6孔板加入含蛋白酶抑制剂、磷酸蛋白酶抑制剂的RIPA裂解液,冰上裂解4℃,12000 rpm离心10 min。BCA法检测蛋白含量。取30 μg蛋白,进行电泳,再将蛋白电转移至PVDF膜上,室温封闭1 h。加入一抗(1∶1000)4℃孵育过夜。洗膜5次后室温孵育二抗(1∶5000)。洗膜,ECL发光液显色,天能系列全自动化学发光成像分析系统扫描成像。以Tubulin作为内参,目的蛋白条带与内参比较作为条带的相对表达值。
1.8 统计学处理
采用SPSS21.0统计软件对数据进行分析,重复3次实验数据采用±s表示,两组间比较采用Student'st检测,正态分布的计量资料多组间比较采用单因素方差分析,以P<0.05为差异有统计学意义。
2 结果
2.1 CoCl2诱导C2C12骨骼肌细胞萎缩
正常组可见长条状肌管形成,CoCl2处理后肌管萎缩、断裂(图1)。
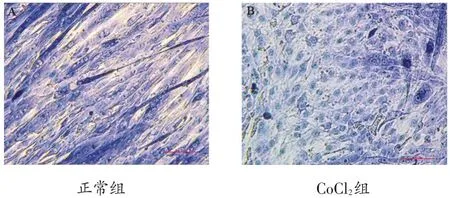
图1 两组形态学比较(×40,bar=100μm)
2.2 CoCl2诱导C2C12骨骼肌细胞ROS含量
使用荧光标记的 ROS分子探针孵育细胞,检测结果(图2)显示:与正常对照组(0.999±0.287)比较,200 μM CoCl2处理C2C12细胞24小时后,荧光强度明显增强(1.833±0.043),差异有统计学意义(t=-4.965,P=0.008)。

图2 两组ROS的含量比较
2.3 CoCl2诱导自噬体形成
正常组具有清晰的双层膜结构的线粒体,呈梭形或者椭圆形,偶见自噬体。CoCl2处理C2C12细胞24小时后,可见空泡化结构,具有清晰双层膜结构的自噬体及髓鞘样结构(图3)。

图3 两组自噬体的形成情况(×10000,bar=0.5 μm)
2.4 CoCl2诱导自噬相关基因mRNA表达
使用CoCl2可诱导BNIP3、LC3B mRNA表达显著增加。与正常组比较,CoCl2组BNIP3 mRNA相对表达量为8.843 ± 0.202,t=-67.239,P=0.000;LC3B mRNA相对表达量为1.348 ± 0.213,t=-2.836,P=0.048(见表1)。HIF-1α mRNA表达与正常组比较无统计学差异(P>0.05)。
表1 两组HIF-1α、BNIP3、LC3B mRNA表达比较(± s)

表1 两组HIF-1α、BNIP3、LC3B mRNA表达比较(± s)
注:与正常组比较,*P<0.05,**P<0.01。
组别正常组CoCl2组t值P值LC3B 1.000±0.000 1.348±0.213*-2.836 0.048 HIF-1α 1.000±0.000 0.816±0.202 1.583 0.188 BNIP3 1.000±0.000 8.843±0.202**-67.239 0.000
2.5 CoCl2诱导自噬相关蛋白表达
使用CoCl2可诱导HIF-1α、BNIP3、LC3蛋白表达显著增加。与正常组比较,CoCl2组HIF-1α蛋白表达为(14768.720 ± 2045.420)%,t=-12.421,P<0.01;BNIP3蛋白表达为(34181.006 ± 3130.082)%,t=-18.859,P<0.01;LC3B蛋白表达为(555.642 ± 66.625)%,t=-11.845,P<0.01(见表2)。
表2 两组HIF-1α、BNIP3、LC3蛋白表达比较(± s)

表2 两组HIF-1α、BNIP3、LC3蛋白表达比较(± s)
注:与正常组比较,*P<0.05,**P<0.01。
组别正常组CoCl2组t值P值HIF-1α(%)100.000±0.000 14768.720±2045.420*-12.421 0.000 BNIP3(%)100.000±0.000 34181.006±3130.082*-18.859 0.000 LC3Ⅱ/LC3Ⅰ(%)100.000±0.000 555.642±66.625**-11.845 0.000
2.6 抑制CoCl2诱导的自噬对MAFbx蛋白表达的影响
使用CoCl2处理24 h后,可观察到MAFbx蛋白表达明显上升。与正常组比较,CoCl2组MAFbx蛋白表达约为正常组的2倍。加入3MA后,MAFbx蛋白表达较CoCl2处理组下降。各组间差异具有统计学意义(F=18.246,P<0.01)(图4)。
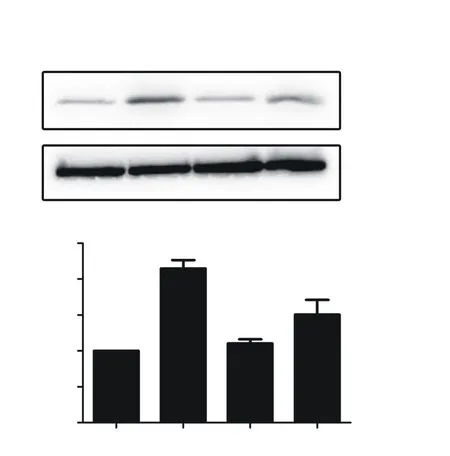
A:Western blot检测MAFbx及内参结果;B:灰度值表示MAFbx相对蛋白表达量;与其它3组比较,**P<0.01。图4抑制自噬后MAFbx蛋白表达变化
3 讨论
缺氧可使骨骼肌萎缩,功能下降[5]。为了阐明低氧环境下肌萎缩机制,本研究采用CoCl2体外模拟低氧环境,观察C2C12小鼠成肌细胞系变化。CoCl2模拟缺氧是国际上比较常用的模拟缺氧的方法,钴离子能竞争血红蛋白卟啉环中的铁离子,抑制氧合血红蛋白的形成而丧失与O2结合的能力,诱发一系列类似缺氧的反应,从而模拟缺氧[7]。在细胞中,CoCl2抑制脯氨酰羟化酶的羟化作用,从而使细胞处于缺氧状态[8]。当机体处于缺氧环境时,线粒体电子传递链中氧分子供应不足,导致大量的ROS产生,产生的ROS会导致细胞形态功能紊乱或细胞死亡[9]。本研究结果显示低氧环境下ROS浓度显著提高。有文献报道细胞内 ROS产生是诱发自噬的主要原因[10]。
本研究中,CoCl2诱导缺氧后可观察到双层膜结构的自噬体形成,LC3Ⅱ/LC3Ⅰ比例上调,表明自噬参与了CoCl2诱导的骨骼肌缺氧。自噬是近年研究的热点之一,是细胞应激过程中产生的一种适应性的代谢活动。自噬形成时,胞浆型LC3(即LC3-Ⅰ)会酶解掉一小段多肽,转变为(自噬体)膜型(即LC3-Ⅱ),因此,LC3-Ⅱ/Ⅰ比值的大小可估计自噬水平的高低[11]。另外,电镜亦作为观察自噬的金标准。自噬的激活受多通路调控,如哺乳类动物雷帕霉素靶蛋白(mammalian target of rapamycin,m-TOR),AMP依赖的蛋白激酶[Adenosine 5‘ -monophosphate(AMP)-activated protein kinase,AMPK]等[12,13]。为进一步研究 CoCl2诱导自噬过程中信号通路的改变,本研究检测了HIF-1α及其下游BNIP3 mRNA及蛋白的表达。HIF-1α是机体维持氧稳态信号系统中最为关键的异二聚体转录因子,其表达受氧浓度影响[14]。在常氧条件下,HIF-1α可被脯氨酸羟化酶羟化,羟基化的HIF-1α通过泛素-蛋白酶体系统迅速降解。低氧环境下,羟基化被抑制,HIF-1α降解受阻,从而激活下游靶基因的表达[15]。BNIP3是BH3-only家族中的一员,是HIF-1α的靶基因之一,在低氧时能被HIF-1α转录激活,表达后定位于线粒体,进一步激活线粒体自噬[16]。有文献报道,HIF-1α/BNIP3参与了心肌缺血再灌注中的自噬过程。抑制BNIP3可减少缺氧引起的心肌细胞死亡,改善心功能[17,18]。另有文献报道,LC3可与BNIP3二聚体上的LC3结合区域(LC3-interacting region,LIR)结合诱导自噬[19,20]。本研究结果显示:HIF-1α、BNIP3、LC3表达上升,提示着HIF-1α/BNIP3参与了骨骼肌缺氧诱导的自噬。但BNIP3是否与LC3直接作用引起自噬需要进一步研究。
MAFbx是肌肉特异表达的泛素E3连接酶,被认为是骨骼肌萎缩的标记物[21]。Pomies等[22]用H2O2处理C2C12肌管18小时后,MAFbx基因表达增强,肌蛋白降解增加,从而导致肌萎缩,运用抗坏血酸处理后可减少ROS的产生及MAFbx的表达,肌管直径变大。肌肉萎缩主要通过两条蛋白降解途径,一为泛素化介导的蛋白降解途径,另一条为自噬-溶酶体途径,但两者间的关系尚有争议[4]。有报道在急性间歇性缺氧(Acute intermittent hypoxia,AIH)大鼠模型中MAFbx表达增加,自噬相关基因LC3和γ-氨基丁酸受体相关蛋白1(Gamma-aminobutyric acid receptor-associated protein-like 1,GABARAPL1)基因mRNA表达上升[23]。另一报道则认为LC3蛋白及自噬相关通路蛋白BNIP3及GABARAPL1与MAFbx表达并无直接关系[24]。本研究使用自噬抑制剂3MA,研究MAFbx与CoCl2诱导自噬间的关系。结果显示,3MA与CoCl2共处理组MAFbx表达较单独CoCl2处理减少,提示抑制自噬可减少MAFbx的表达。
综上,CoCl2诱导C2C12骨骼肌细胞萎缩,可能与HIF-1α/BNIP3信号通路诱导的自噬发生有关,抑制缺氧条件下的自噬可部分减少肌萎缩。
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2022年8期)2022-12-07
保健与生活(2022年13期)2022-07-06
体育科技文献通报(2022年3期)2022-05-23
医学综述(2021年16期)2021-12-01
疯狂英语·新阅版(2021年8期)2021-09-10
天津医科大学学报(2021年3期)2021-07-21
中国特种设备安全(2019年4期)2019-05-20
北京广播电视报(2019年8期)2019-03-27
武警医学(2019年2期)2019-03-05
运动(2018年14期)2018-07-16
