
Electroacupuncture exerts neuroprotective effects on ischemia/reperfusion injury in JNK knockout mice: the underlying mechanism
2018-08-09 01:27:30ChunXiaoWuYiHuiFengLuYangZhuLianZhanXiuHongXuXiaoYingHuZhiHuaZhuGuoPingZhou
中国神经再生研究(英文版) 2018年9期
Chun-Xiao Wu , Yi-Hui Feng , Lu Yang , Zhu-Lian Zhan, Xiu-Hong Xu, Xiao-Ying Hu, Zhi-Hua Zhu, Guo-Ping Zhou ,
1 School of Traditional Chinese Medicine, Southern Medical University, Guangzhou, Guangdong Province, China
2 Medical College of Acu-Moxi and Rehabilitation, Guangzhou University of Chinese Medicine, Guangzhou, Guangdong Province, China
3 Central Hospital of Qingdao, Qingdao, Shandong Province, China
4 Traditional Chinese Medicine-Integrated Hospital, the Affiliated Hospital of Southern Medical University, Guangzhou, Guangdong Province, China
5 Ruikang Hospital Affiliated to Guangxi College of Traditional Chinese Medicine, Nanning, Guangxi Zhuang Autonomous Region, China
Abstract Simple regulation of c-Jun N-terminal kinase (JNK) or p38 mitogen-activated protein kinase (MAPK) pathways is not enough to trigger cell apoptosis. However, activation of the stress activated pathway (JNK/p38 MAPK) together with inhibition of the growth factor activated extracellular signal-regulated kinase (ERK) pathway can promote cell apoptosis. We hypothesized that inhibition of the JNK or p38 pro-apoptotic pathway and activating the ERK pathway could be the mechanism of anti-apoptosis following cerebral ischemia/reperfusion injury. To investigate the mechanism of the protective effect of electroacupuncture on cerebral ischemia/reperfusion injury in JNK knockout mice, mouse models of cerebral ischemia/reperfusion injury were established by Longa’s method. Electroacupuncture was conducted at acupoints Chize(LU5), Hegu (LI4), Sanyinjiao (SP6) and Zusanli (ST36) 1.5 hours after ischemia/reperfusion injury for 20 minutes, once a day. The neurological function was evaluated using neurological deficit scores. The expression of phospho-extracellular signal-regulated kinase (p-ERK) and phospho-p38 (p-p38) in JNK knockout mice was detected using double-labeling immunofluorescence and western blot assay. The mRNA expression of ERK and p38 was measured by quantitative real-time polymerase chain reaction. Electroacupuncture improved neurological function, increased the immunoreactivity and relative expression of p-ERK and reduced that of p-p38 in the cerebral cortex and hippocampus on the injured side. Electroacupuncture increased mRNA expression of ERK, but decreased that of p38 in the cerebral cortex and hippocampus on the injured side. In conclusion, electroacupuncture upregulated the protective ERK pathway and inhibited the pro-apoptotic p38 pathway,thereby exerting a neuroprotective effect and improving the neurological function in JNK knockout mice.
Key Words: nerve regeneration; MAPKs signaling pathway; cerebral ischemia/reperfusion; stress-activated pathway; growth factor-activated pathway; electroacupuncture; apoptosis
Introduction
Cerebral ischemia leads to inflammation, oxidative damage and programmed cell death that aggravate brain tissue damage and seriously influence the daily life of stroke patients (Dirnagl,2012). The mechanism of cerebral ischemia/reperfusion (I/R)injury is mainly associated with inflammation and apoptosis(Broughton et al., 2009; Eltzschig and Eckle, 2011). Reducing apoptosis helps the recovery of brain functions (Bhuiyan and Fukunaga, 2009).
Mitogen-activated protein kinases (MAPKs) play a role in neural cell growth and proliferation (Irving and Bamford,2002). The MAPKs signaling pathway is widely expressed in the nervous system and regulates the repair or apoptosis of nerve cells after cerebral I/R injury (Abe et al., 2000; Shaw and Kirshenbaum, 2006; Kovalska et al., 2012). The main three subfamilies of MAPKs; extracellular signal-regulated kinases(ERKs), c-Jun N-terminal kinases (JNKs), and p38 kinases,are involved in regulating dysfunction and apoptosis of neuronal cells after cerebral I/R injury (Yang et al., 2003; Hu et al.,2012). The ERK signaling pathway starts the self-repair process, which plays an important role in recovering from cerebral I/R injury (Shioda et al., 2009). p38 and JNK serve as mediators of cellular stresses, which are involved in inflammation and neuronal cell apoptosis (Yoshida et al., 2010; Kyriakis and Avruch, 2012). A previous study also proved that cell survival or apoptosis was dependent on the dynamic balance of the growth factor-activated (ERK pathway) and stress-activated(JNK/P38 signaling pathway) pathways (Junttila et al., 2008).Broad “crosstalk” between several signaling pathways has been reported and could lead to mutual synergy or inhibition between pathways (Junttila et al., 2008). Another study suggested that simple JNK or p38 MAPK was not enough to trigger the process of apoptosis. Only the concurrent activation of stress-activated pathway (JNK/p38 MAPK) coupled with the restraint of the growth factor-activated ERK pathway could promote cell apoptosis (Xia et al., 1995). We propose that inhibiting the JNK or p38 pro-apoptotic pathway and simultaneously activating the ERK protective pathway could result in an anti-apoptotic mechanism after cerebral I/R injury. Therefore,we knocked out one of the stress-activated pathways (JNK)and investigated the effect of electroacupuncture (EA) on ERK and p38 kinase pathways in the JNK knockout mice after I/R injury.
EA therapy is advantageous in improving neurological deficit symptoms and rehabilitating limb motor function after cerebral I/R injury (Chavez et al., 2017). “Chize (LU5), Hegu(LI4)”, “Sanyinjiao (SP6) and Zusanli (ST36)” served as common acupoints for the clinical treatment of stroke (Xiong et al., 2008; Yang et al., 2015). A previous study found that EA could reduce the area of cerebral infarction and neuronal apoptosis in JNK knockout mice with cerebral I/R injury (Feng,2015). However, the role of EA in the anti-apoptotic mechanism of JNK knockout mice with cerebral I/R injury needs further investigation. This study aims to elucidate the possible mechanisms of EA action in JNK knockout mice with cerebral I/R injury.
Materials and Methods
Animals
A total of 54 JNK knockout mice (half males and half females),weighing 20–25 g, were purchased from Cyagen Biosciences Co. Ltd. (Guangzhou, China; license number: SCXK (Yue)2013-0002). The experimental procedure followed the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1986), and was approved by the Animal Ethics and Welfare Committee of Southern Medical University of China (approval No. 2013-032). The mice were fed standard rodent chow, and allowed free access to water. The mice were acclimatized to the animal house that was maintained at a room temperature between 20° C and 22° C and a relative humidity of 65–70%. The JNK knockout mice were randomized into sham group (sham control), I/R group (model), and EA group (I/R + EA) (n =18 per group).
Establishing cerebral I/R injury mouse model
The I/R injury model was established by middle cerebral artery occlusion, as described previously (Longa et al., 1989). Briefly, after anesthetizing the mice with 10% chloral hydrate (300 mg/kg) by intraperitoneal injection, the left common carotid artery, left internal carotid artery, external carotid artery, and vagus nerve were carefully exposed via a midline incision under a surgical microscope. Approximately 11 ± 0.5 mm of a nylon suture was introduced into the internal carotid artery to occlude the middle cerebral artery until a slight resistance was observed during insertion. Reperfusion was achieved by withdrawing the thread slowly to restore the blood supply to the ischemic area after 30 minutes of occlusion. The mice in the sham group underwent the same surgical procedures as described earlier but arterial occlusion was not performed.
Electroacupuncture intervention
The mice in the EA group underwent EA stimulation 1.5 hours after I/R injury (then once a day until sacrifice). Stainless steel acupuncture needles (diameter: 0.16 × 13 mm2; Suzhou Universal Acupuncture Medical Devices Co., Ltd., Suzhou,China) were inserted 2–3 mm into LU5, LI4, ST36 and SP6 acupoints of the paralyzed limb. They were located according to the Experimental Acupuncture Science edited by Li (2007).The location of the selected acupoints were as follows: LU5,in the depression of outer end of the transverse cubital crease;LI4, between 1stmetacarpal bone and 2ndmetacarpal bone ;ST36, 3.5 mm below the fibular head at the outer lateral posterior knee; SP6, 5 mm above the tip of the inner ankle of the posterior limb. The acupoints were stimulated for 20 minutes with a dilatational wave of frequency 5/10 Hz and intensity 2 mA using an EA instrument (Model KWD-808I, Suzhou Universal Acupuncture Medical Devices Co., Ltd.).
Neurological deficit scores
The neurological deficit scores were evaluated in six mice at 2 hours, 1 and 3 days after the operation using a 5-point neurological scale according to a previous method (Longa et al.,1989). The neurological deficit scores were defined as follows:score 0, no observable deficit; score 1, failure to fully extend the right forepaw; score 2, circling to the contralateral side; score 3,falling to the contralateral side; and score 4, loss of walking or consciousness. The mice with scores 0 or 4 were excluded.
Tissue sampling
After terminal anesthesia, rapid perfusion with 0.9% normal saline (NaCl) and 4% paraformaldehyde in phosphate buffered saline for 3–5 minutes eliminated the influence of blood factors. The whole brain were dissected out of the cranial cavity quickly and washed with ice-cold saline. The separated brain tissue was frozen in liquid nitrogen and stored at −80 ° C pending subsequent double-labeling immunofluorescence,quantitative real-time polymerase chain reaction (PCR) and western blot assay.
Double-labeling immunofluorescence
The expression of phospho-ERK (p-ERK) and phospho-p38(p-p38) on the injured side of the cortex and hippocampus at 2 hours, 1 day and 3 days (Shipp, 2007) were measured using the double-labeling immunofluorescence in six mice at each time point. Frozen sections were boiled in EDTA antigen repair solution (pH 8.0) (Cell Signaling Technology, Danvers,MA, USA) for antigen retrieval and blocked with 3% bovine serum albumin for 30 minutes at room temperature. For double-labeling immunofluorescence, the sections were incubated with a primary antibody against p-p38 monoclonal antibody(anti-rabbit, 1:100; Cell Signaling Technology, Boston, USA)diluted in phosphate-buffered saline at 4°C overnight. After washing, the sections were incubated with the corresponding secondary antibody (cy3-goat anti-rabbit, 1:300; Google Biological Technology Co., Ltd., Wuhan, China). Next, the sections were incubated with an anti-p-ERK primary monoclonal antibody (anti-mouse, 1:100; Abcam, Cambridge, UK) and a secondary antibody (488 goat anti-mouse, 1:400; Google Biological Technology Co., Ltd.). The sections were observed and imaged under a laser scanning confocal microscope (LSM-510META; Zeiss, Jena, Germany). Positive integrated optical densities (IODs) of p-ERK or p-p38 were evaluated using the Image-Pro Plus 6.0 software (Media Cybernetics, Bethesda,MD, USA). IOD values = Positive area in immunofluorescence image × optical density. A higher IOD value corresponded to a higher expression of p-ERK, or p-p38.
Quantitative real-time PCR
Quantitative real-time PCR was performed to measure the mRNA expression of ERK and p38 in the injured brain in three samples in each group. The total RNA was extracted from the left (injured) side of brain at 2 hours, 1 day and 3 days using the TRIzol reagent (Life Technologies, Carlsbad, CA, USA).Afterwards, 2.0 µg of total RNA was reverse-transcribed to cDNA using a Toyobo First Strand cDNA Synthesis Kit (Qiagen, Hilden, Germany). Quantitative real-time PCR was performed using a THUNDERBIRD SYBR q-PCRMix (Toyobo,Osaka, Japan). The PCR primer sequence was synthesized by the Invitrogen Company (Shanghai, China) as follows: β-actin: forward 5′-GTG ACG TTG ACA TCC GTA AAG A-3′,reverse 5′-GTA ACA GTC CGC CTA GAA GCA C-3′; ERK:forward 5′-TGG AGA CGG ACC TTT ACA AGC-3′, reverse 5′-CAA GTG GTG TTC AGC AGG AGG-3′; and p38:forward 5′-GAC CTA CTG GAG AAG ATG CTC GTT-3′,reverse 5′-TTT CAA AGG ACT GGT CAT AAG GGT-3′.cDNA was amplified under the following cyclic conditions:95° C for 10 minutes, followed by 40 cycles of 95° C for 15 seconds and 60°C for 60 seconds. The mRNA expression of ERK and p38 was normalized to the expression level of β-actin and calculated using the following equation: fold change =2–ΔΔCT.
Western blot assay
Proteins from the brain tissue of three samples in each group,3 days after I/R, were extracted and homogenized in a radioimmunoprecipitation assay lysis buffer. The proteins were electrophoresed using 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (Millipore, Boston, USA). The membranes were blocked with 5% nonfat dry milk for 1 hour and incubated with rabbit anti-ERK polyclonal antibody (1:1000;Affinity Biosciences, OH, USA), p-ERK polyclonal antibody(1:1000; Affinity Biosciences), p38 monoclonal antibody(Abcam), p-p38 monoclonal antibody (1:1000; Cell Signaling Technology, Boston, MA, USA), and mouse anti-glyceraldehyde-3-phosphate dehydrogenase monoclonal antibody(1:1000; Google Biological Technology Co., Ltd.) at 4°C overnight. The membranes were incubated with a corresponding secondary antibody (1:3000; goat anti-rabbit/mouse IgG;Google Biological Technology Co., Ltd.) in Tris-buffered saline and Tween 20 for 30 minutes. The blots were developed using an enhanced chemiluminescence, and the intensity of the bands was measured using the AlphaEaseFC analyzer software(AlphaInnotech, San Leandro, CA, USA). The optical density value ratio of the target band to the internal reference served as the relative expression of the target protein.
Statistical analysis
All data, presented as the mean ± SD, were analyzed using SPSS 21.0 software (SPSS, Chicago, IL, USA). The statistical analysis of the data was conducted using the one-way analysis of variance via least significant difference post hoc comparison when comparing more than two groups. If the tested data were not normally distributed, the nonparametric test was adopted.A P value less than 0.05 was considered statistically significant.
Results
Effects of EA on the neurological function of JNK knockout mice with I/R injury
The neurological deficit scores were evaluated in the sham, I/R,and EA groups. The mice in the I/R and EA groups presented with obvious neurological deficits by two hours after I/R, while the JNK knockout mice in the sham group did not manifest any signs of cerebral damage. Figure 1 shows that there were no significant differences between the neurological deficit scores of the I/R and EA groups at 2 hours and 1 day after I/R injury(P > 0.05). However, the neurological deficit scores of the EA group were significantly lower than those of the I/R group 3 days after I/R injury (P < 0.05).
Effects of EA on the p-ERK and p-p38 immunoreactivities in the hippocampus and cortex of the JNK knockout mice with I/R injury
The expression of p-ERK and p-p38 signaling pathways in the JNK knockout mice with I/R injury was detected using double-labeling immunofluorescence (Figure 2C, F). Figure 2A,B show that both p-ERK and p-p38 immunoreactivities in the hippocampus increased to a peak at 1 day after I/R injury, and stabilized at 3 days. EA increased the p-ERK immunoreactivity in the JNK knockout mice with I/R injury at 1 day and 3 days(P < 0.05). However, the p-p38 immunoreactivity was lower in the EA group than in the I/R group at 1 day and 3 days (P <0.05). Similar results were found in the cortex (Figure 2D, E).
Effects of EA on the mRNA expression levels of ERK and p38 in the JNK knockout mice with I/R injury
To investigate further the mechanism underlying the protective effect of EA treatment on the expression of ERK and p38,the mRNA expressions of ERK and p38 were evaluated in the JNK knockout mice using quantitative real-time PCR. Figure 3 shows that the mRNA expressions of ERK and p38 were upregulated, peaking at 1 day, then decreasing by 3 days. However, the mRNA expressions of ERK and p38 were not significantly different between the I/R and EA groups at the 2-hour time point (both P > 0.05). After 1 and 3 days of I/R injury,the mRNA expression of ERK was enhanced and that of p38 diminished in the EA group compared with the I/R group (all P < 0.05; Figure 3).
Effects of EA on the protein expression of p-ERK and p-P38 in the JNK knockout mice with I/R injury
Western blot assay results showed the target protein expression of p-ERK and p-p38 at 3 days post I/R. Compared with the I/R group, the protein expression of p-ERK was upregulated and that of p-p38 was downregulated in the EA group (both P< 0.05), which was consistent with the finding of double-labeling immunofluorescence. However, the protein expression of p-ERK in the sham group was similar to that in the I/R group,and the protein expression of p-p38 in the EA group was not significantly different from that in the sham group (P > 0.05;Figure 4).
Discussion
Cerebral ischemia involves a series of pathological changes,such as inflammation, oxygen deficit, brain edema and nerve cell apoptosis, leading to brain injury and reduced limb motor function. Reperfusion causes further brain injury and apoptosis. The neurological deficit symptoms we observed indicated neurological dysfunction and brain damage following I/R injury (Vaibhav et al., 2013; Yaidikar and Thakur, 2015). We found that EA could alleviate the neurological deficits in JNK knockout mice with cerebral I/R injury, especially after 3 days.Other studies have also indicated that EA could improve the limb locomotor function and provide some neuroprotective effects against I/R injury (Zhang et al., 2014b; Lu et al., 2015;Liu et al., 2016). EA has been shown to reduce cerebral infarct areas and inhibit apoptosis of nerve cells in the JNK knockout mice with I/R injury (Feng, 2015).
The possible mechanisms underlying the neuroprotective effects of EA against I/R injury may be associated with the regulation of MAPK family. The MAPK family plays an important role in I/R injury; it regulates neural cell death and survival via signaling pathways related to cell apoptosis (Nozaki et al.,2001). The MAPK family mainly consists of ERK, p38, and JNK signaling pathways. Although the JNK signaling pathway had been knocked out in our study, double-labeling immunofluorescence showed that the expression of p-ERK was still upregulated and the expression of p-p38 was downregulated in the EA group compared with the I/R group at the same time points. These results indicate that EA might inhibit neuronal apoptosis and improve neurological deficit symptoms by activating the expression of p-ERK and inhibiting the expression of p-p38. Our western blot assay and quantitative PCR data further verified the double-labeling immunofluorescence findings. The MAPK/ERK pathway is mainly involved in regulating cell proliferation, migration, differentiation, and reducing apoptosis (Sun et al., 2015), whereas the activation of p38 promotes neuronal apoptosis (Kim and Choi, 2015). The ERK activated by phosphorylation can inhibit caspase-9 processing and activation, blocking the caspase cascade and inhibit apoptosis (Allan et al., 2003). A related study has indicated that the inhibition of ERK activation by ERK inhibitors in vitro and in vivo can weaken the expression of cleaved caspase-3 to protect neural cells from brain injury-induced apoptosis (Zhao et al., 2012). Previous studies have also found that EA can inhibit neural cell apoptosis during I/R injury by activating the ERK signaling pathway (Cheng et al., 2014b; Wu et al., 2015), which is consistent with the present findings. However, p38 mediates inflammation and promotes apoptosis.
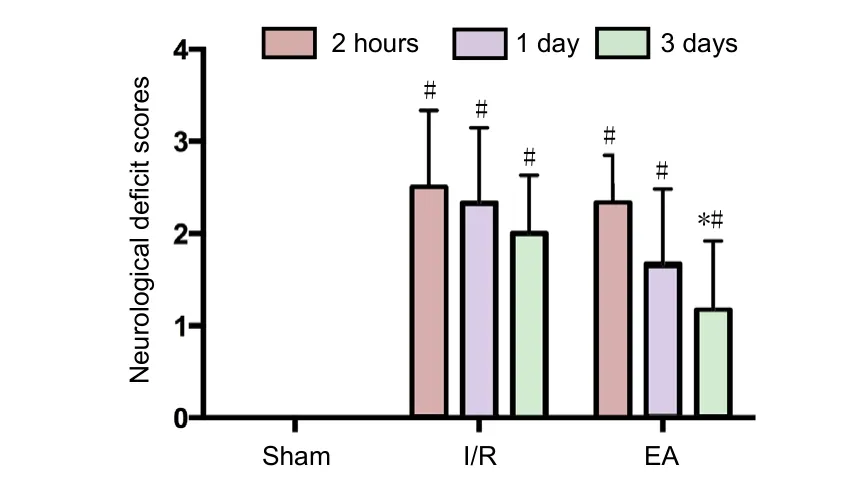
Figure 1 Effects of Electroacupuncture on the neurological deficit of the JNK knockout mice with I/R injury.
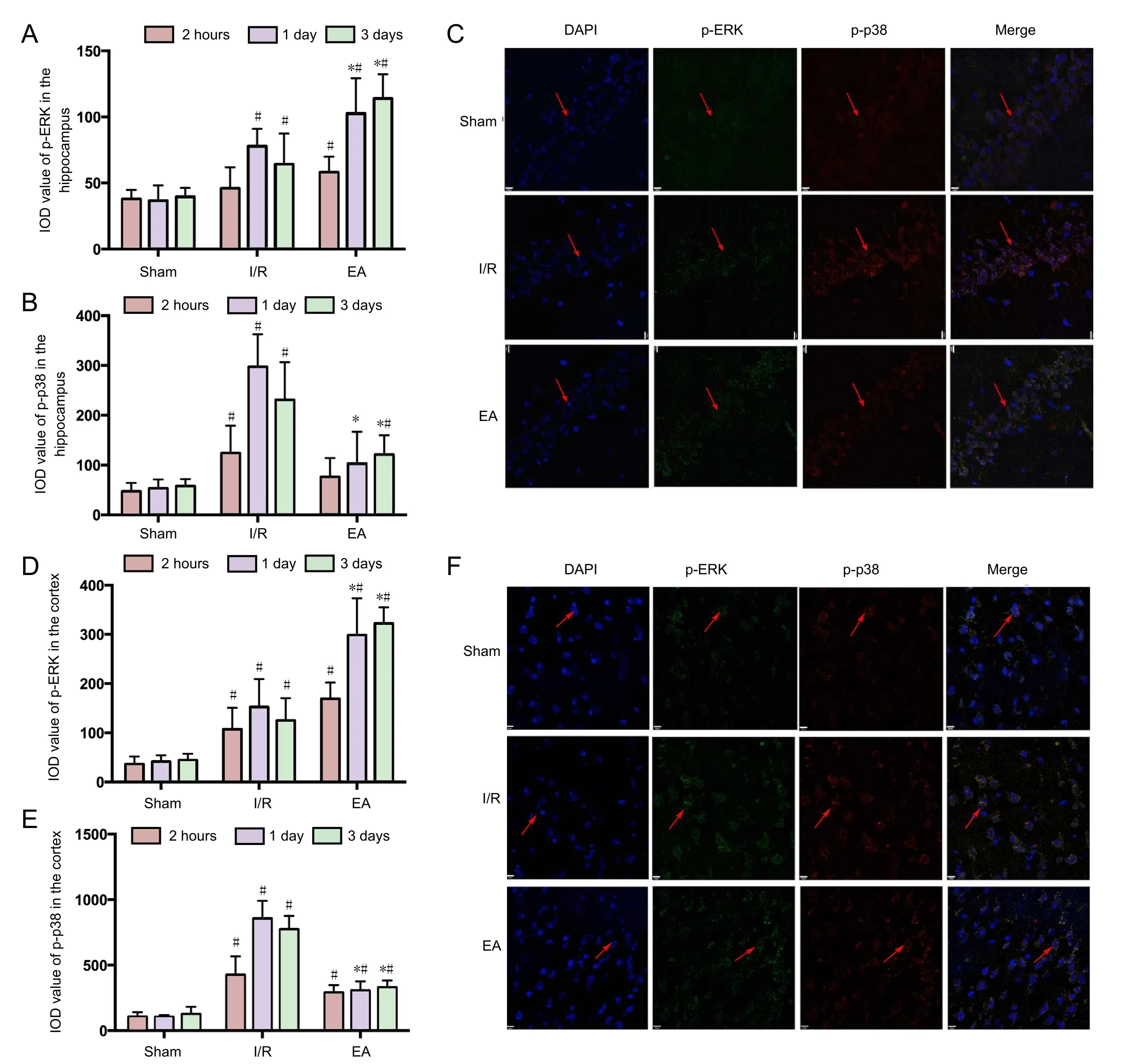
Figure 2 The p-ERK and p-p38 immunoreactivity in the hippocampus and cortex of JNK knockout mice with cerebral I/R injury .
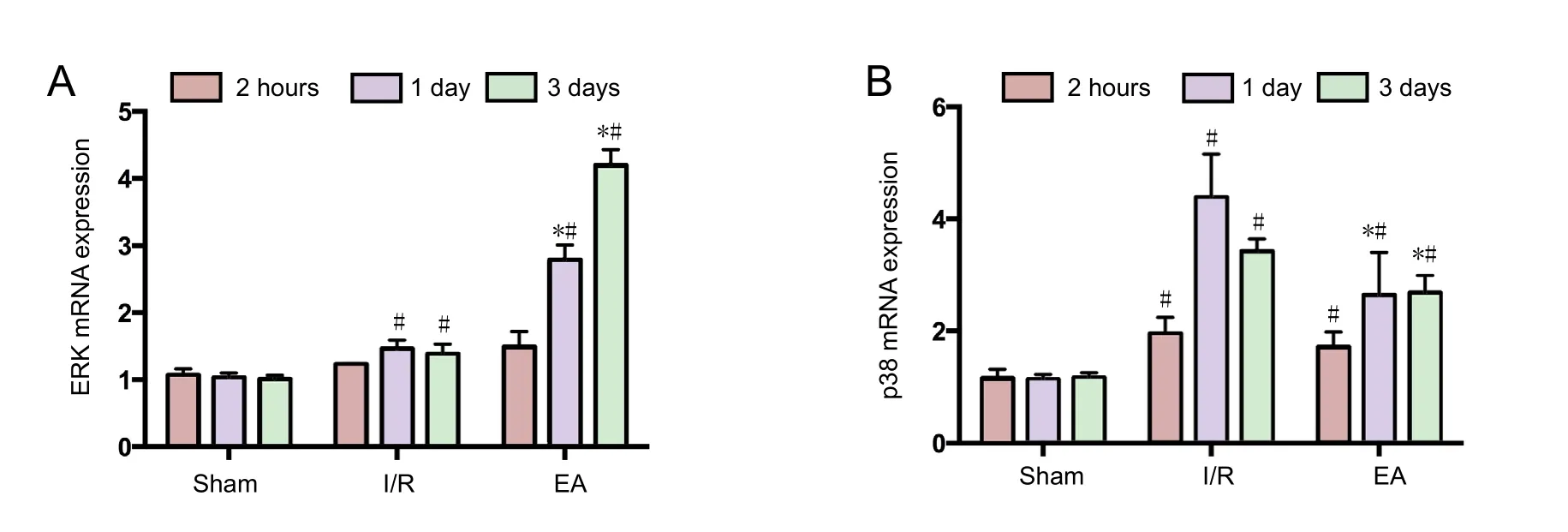
Figure 3 The mRNA expression of ERK and p38 in the injured brain tissues of JNK knockout mice with cerebral I/R injury.
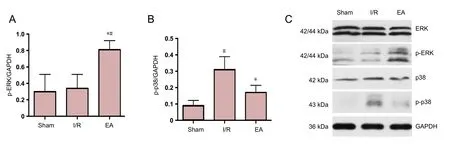
Figure 4 Protein expression of p-ERK and p-P38 in the JNK knockout mice with I/R injury.
Studies have shown that the activation of p38 induces further downstream transcription factors and activates caspase-3,caspase-6, and caspase-7, thereby activating the genes associated with apoptosis, resulting in nerve cell apoptosis (Morel et al., 2005; Chan et al., 2009). Inhibition of p38 phosphorylation can promote the phosphorylation of ERK1/2, alleviate brain injury through restraining autophagic cell death and rescue degenerative neural cells in ischemic penumbra (Zhang et al.,2014a). Cheng et al. (2014a) reported that EA at acupoints can inhibit the expression of p38, thereby reducing the oxidative/nitrative stress and downregulating the tumor necrosis factor-α/TRADD/FADD/cleaved caspase-8/cleaved caspase-3 apoptotic pathway to attenuate neural cell apoptosis.
The findings of previous studies and the present study suggest that EA stimulation exerts an anti-apoptotic effect against I/R injury, which involves upregulating the expression of p-ERK and inhibiting the expression of p-p38 at the same time.The p-ERK signaling pathway was significantly upregulated and the p-p38 signaling pathway was downregulated in the same brain section from the EA group compared with a similar section from the I/R group, and at both 1 and 3 days. This indicates that EA could synchronously regulate ERK and p38 signaling pathways in the JNK knockout mice with I/R injury.It confirms the previous assumption that EA treatment exerts a neuroprotective effect against cerebral I/R injury, which may not simply rely on one of the MAPK signaling pathways, but requires simultaneous regulation of several MAPK signaling pathways. EA may regulate cerebral I/R injury in JNK knockout mice by activating the neuroprotective pathway (ERK) and inhibiting the p38 pro-apoptotic pathway (MAPK signaling pathway). This suggests some “crosstalk” exists between ERK and p38. Other research has indicated that inhibiting the expression of p38 might stimulate the phosphorylation of ERK signaling pathway (Lee et al., 2002; Li et al., 2003), and that the activation of p38/JNK might shut off the ERK pathway-mediated survival signaling and induce apoptosis (Junttila et al.,2008). Therefore, EA may upregulate the expression of p-ERK and downregulate the expression of p-p38 at the same time to promote the crosstalk between the two signaling pathways that can exert a better neuroprotective effect against cerebral I/R injury.
Early EA intervention after cerebral ischemia can increase cerebral blood flow, reduce the extent of cerebral I/R injury,and expand the therapeutic time window (Ren et al., 2010; Kim et al., 2013). This indicated that the best time for acupuncture treatment is within 3 hours of ischemia (Wang et al., 2012).We chose 1.5 hours after cerebral I/R injury for EA treatment.The double-labeling immunofluorescence and quantitative real-time PCR results indicated that the expression of ERK was upregulated and the p38 expression was suppressed at 1 and 3 days in the EA group compared with the I/R group. However,no significant difference was found between the EA and I/R groups at 2 hours. Nerve cell apoptosis usually reaches a peak at 24–48 hours after cerebral I/R injury (Zhang and Li, 2012).The EA-regulated ERK and p38 signaling pathways in JNK knockout mice with cerebral I/R injury were obvious 1 day after cerebral I/R injury, which was the peak point of apoptosis.The aftereffects of EA and self-repair mechanisms during the increase in apoptosis would promote the ERK protective pathway and inhibit the p38 pro-apoptotic pathway simultaneously to exert a neuroprotective effect. Therefore, EA would not have an immediate effect after I/R injury in the JNK knockout mice. Rather, EA can exert a neuroprotective effect 1 day after I/R injury compared with the I/R group. A significant difference was found between the EA and I/R groups after 3 days,indicating that EA intervention once a day continued to exert an effect and regulate the expression of ERK and p38 in JNK knockout mice to alleviate cerebral I/R injury.
However, this study still has limitations. First, the I/R injury model was established by middle cerebral artery occlusion,which may have a high death rate and model establishment takes a long time. A better model for I/R injury established by an optimized method is needed in future research. Second,we used double-labeling immunofluorescence to detect the expression of p-ERK and p-p38. The crosstalk between antibodies may affect the expression of the target pathway to some extent. Therefore, a comprehensive method with less interference is needed to measure the expression of p-ERK and p-p38 simultaneously in a future study.
EA at acupoints “LU5,” “LI4,” “ST36,” and“SP6” could alleviate neurological deficit symptoms and reduce apoptosis in the JNK knockout mice with I/R injury.Enhancing the ERK protective pathway and inhibiting the p38 pro-apoptotic pathway simultaneously may be one of the key mechanisms of EA treatment for cerebral I/R injury. EA treatment may be most effective against apoptosis one day after I/R.Three days afterwards maybe a good time point to measure the neuroprotective effect of accumulative acupuncture.
Acknowledgments: The authors are very grateful to all staff from Experimental Animal Center of Southern Medical University of China and thank all the researchers involved in animal experiments.
Author contributions: GPZ obtained funding, participated in study concept and design, and paper authorization. XHX and ZHZ purchased animals and material instruments. LY and CXW analyzed data. CXW wrote the paper. YHF operated the acupuncture. ZLZ and XYH ensured the integrity of the data. All authors approved the final version of the paper.Conflicts of interest: We declare that we have no conflict of interest.
Financial support: This study was supported by the National Natural Science Foundation of China, No. 81173355. The funding body played no role in the study design, in the collection, analysis and interpretation of data, in the writing of the report, or in the decision to submit the article for publication.
Institutional review board statement: All experimental procedures were approved by the Animal Ethics and Welfare Committee of Southern Medical University of China (approval No. 2013-032). The experimental procedure followed the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1985).
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open access statement: This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix,tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers: Clarissa Cavarsan, Federal University of Paraná,Brazil; Lijie Zhai, University of Illinois at Chicago, USA.

- 中国神经再生研究(英文版)的其它文章
- Seeing the wood for the trees: towards improved quantification of glial cells in central nervous system tissue
- 3′-Daidzein sulfonate sodium protects against memory impairment and hippocampal damage caused by chronic cerebral hypoperfusion
- The novel chalcone analog L2H17 protects retinal ganglion cells from oxidative stress-induced apoptosis
- Treatment with NADPH oxidase inhibitor apocynin alleviates diabetic neuropathic pain in rats
- Saikosaponin a increases interleukin-10 expression and inhibits scar formation after sciatic nerve injury
- Various changes in cryopreserved acellular nerve allografts at −80° C