
连作对花生土壤酶活性、养分含量和植株产量的影响①
2018-07-27 07:50江立庚唐荣华郭文峰
土壤 2018年3期
李 忠,江立庚,唐荣华,郭文峰
连作对花生土壤酶活性、养分含量和植株产量的影响①
李 忠1,2,江立庚2,唐荣华1,3,郭文峰1
(1 广西作物遗传改良生物技术重点开放实验室,南宁 530007;2 广西大学农学院,南宁 530007;3 广西农业科学院经济作物研究所,南宁 530004)
通过盆栽试验,设置4个不同连作年限处理,研究连作对花生栽培种桂花17和野生种土壤酶活性、养分含量和植株干物质产量影响的差异。结果表明:与对照相比,两个花生品种不同连作年限土壤有效养分含量和植株干物质产量均降低,相比桂花17下降幅度小。桂花17各连作年限处理土壤的脲酶、转化酶、酸性磷酸酶活性随连作年限的增加有下降的趋势,处理间变幅大。而各生育期土壤的脲酶、转化酶、酸性磷酸酶活性总体表现无一致性上升或下降的规律,处理间变幅小。两个花生品种在不同的连作年限间的过氧化氢酶活性的变化无规律性。连作条件下,花生野生种的土壤理化和生物学性状的变化幅度更小。
花生;土壤酶活性;土壤养分;干物质产量
花生连作可使土壤理化性状恶化,土壤肥力下降,病害增加等,严重的可导致花生产量与品质大幅下降。有关花生栽培种连作障碍的研究已取得一定进展,花生栽培种产生连作障碍的因素主要有土壤酶活性降低,土壤养分不平衡,土壤化感物质累积及土壤致病微生物数量增加等[1-2]。土壤酶及土壤养分一直是研究作物连作障碍的热点,并已在许多作物上报道[3-6]。花生属野生种具有丰富的遗传变异性,也存在很多花生栽培种所不具备的优良性状,对一些病虫害的抗性显著高于花生栽培种[10-11]。花生野生种在其抗病性、遗传差异和杂交利用上的研究已有大量研究报道[7-9],但目前有关花生野生种连作对土壤酶、土壤养分及生物产量的研究还鲜见报道。因此,本研究通过对连作花生根系土壤酶活性、土壤养分及生物产量的测定,初步研究连作条件下花生野生种()及花生栽培种桂花17土壤理化和生物学性状的差异,为鉴定花生野生种的耐连作能力提供依据,为其应用于花生遗传改良提供理论基础。
1 材料与方法
1.1 试验材料
供试花生品种共两个,第一个是花生属花生区组野生种,来自国家种质野生花生南宁分圃第二个为广西主栽花生品种之一:桂花17,由广西农科院经济作物研究所育成和提供。
1.2 盆栽试验及样品采集
供试连作花生2 a土壤及轮作土壤为石灰土,取自于2011年7月广西南宁市双定乡的同一花生种植地块,土壤含速效氮 37.8 mg/kg,有效磷21.8 mg/kg,速效钾36.5 mg/kg,有机质13 g/kg。挖掘其耕作层,一次性基施肥料,氮、磷、钾肥施用量按N 45 kg/hm2,P2O575 kg/hm2,KO245kg/hm2施入,混匀土壤与肥料后装入塑料盆中,塑料盆上口直径35 cm,下口直径25 cm,高25 cm,每盆装土样12 kg,用于种植花生及试验观测。2012年3月分别种植两个花生品种于土壤塑料盆中,塑料盆钵15 cm埋于土中,以一年内连续种植两造花生设计连作处理及种植一造花生闲置一造的休闲方式设计不连作处理,设置连作0(对照CK)、2、4、6 a四个处理(至2014年时实际的连作年限),各处理设计见表1。每个处理种16盆,每盆种3株,3次重复。于花生苗期、开花期、结荚期和成熟期取样,同处理每盆采集10 ~ 20 cm土壤层作为土样,同一土样混匀后,过1 mm筛,用于土壤酶活性和土壤养分含量的测定。

表1 不同连作年限花生土壤处理设计
1.3 土壤酶活性测定
在花生苗期、开花期、结荚期和成熟期进行采样测定,参照关松荫的方法[12]:过氧化氢酶活性采用高锰酸钾滴定法测定,活性以24 h后1 g干土消耗0.1 mol/L KMnO4的体积(ml)表示;脲酶活性采用苯酚-次氯酸钠比色法测定,活性以24 h 后1 g干土中NH3-N 的质量(mg)表示;磷酸酶活性采用磷酸苯二钠法测定,活性以24 h后1 g干土中酚的质量(mg)表示;转化酶活性采用3,5-二硝基水杨酸比色法测定,活性以24 h后1 g干土中葡萄糖的质量(mg)表示。
1.4 土壤养分测定
于花生成熟期时进行采样测定,土壤有机质采用重铬酸钾容量法测定;碱解氮采用碱解扩散法测定;有效磷采用0.5 mol/L碳酸氢钠浸提-钼锑抗比色法测定;速效钾采用1 mol/L乙酸铵提取,火焰光度计法测定[13]。
1.5 植株干物质量测定
在花生苗期、开花期、结荚期和成熟期每处理取8株样本,在 105℃下杀青30 min,之后在75℃烘干至恒重,自然冷却至室温,分别测定其干物质量。
1.6 数据处理
本文采用SPSS19.0软件对试验数据进行统计分析与相关分析,采用Excel软件作图。
2 结果与分析
2.1 花生连作对土壤酶活性的影响
2.1.1 脲酶 由图1可知,桂花17各个处理的土壤脲酶活性总体上苗期处于较低水平,至开花期增加达到最高,至结荚期和成熟期又降低。桂花17各时期各连作年限处理的土壤脲酶活性均比CK低,其中在苗期和开花期各连作年限处理的土壤脲酶活性与CK差异达显著水平(<0.05)。在开花期各连作年限处理的脲酶活性与CK相比下降最多,连作2 a处理比CK下降16.6%,连作4 a处理下降27.8%,连作6 a处理下降25.0%。
而花生各生育期总体表现为,连作2 a和4 a处理土壤脲酶活性降低,连作6 a处理增加,各连作年限处理的土壤脲酶活性总体上升和下降的变化幅度不大。与桂花17相似,在开花期各连作年限处理的土壤脲酶活性均比CK低,连作2 a处理相比CK土壤脲酶活性下降8.3%,连作4 a处理下降13.5%,连作6 a处理下降4.9%,但其他生育期各处理间的土壤脲酶活性差异均不显著。
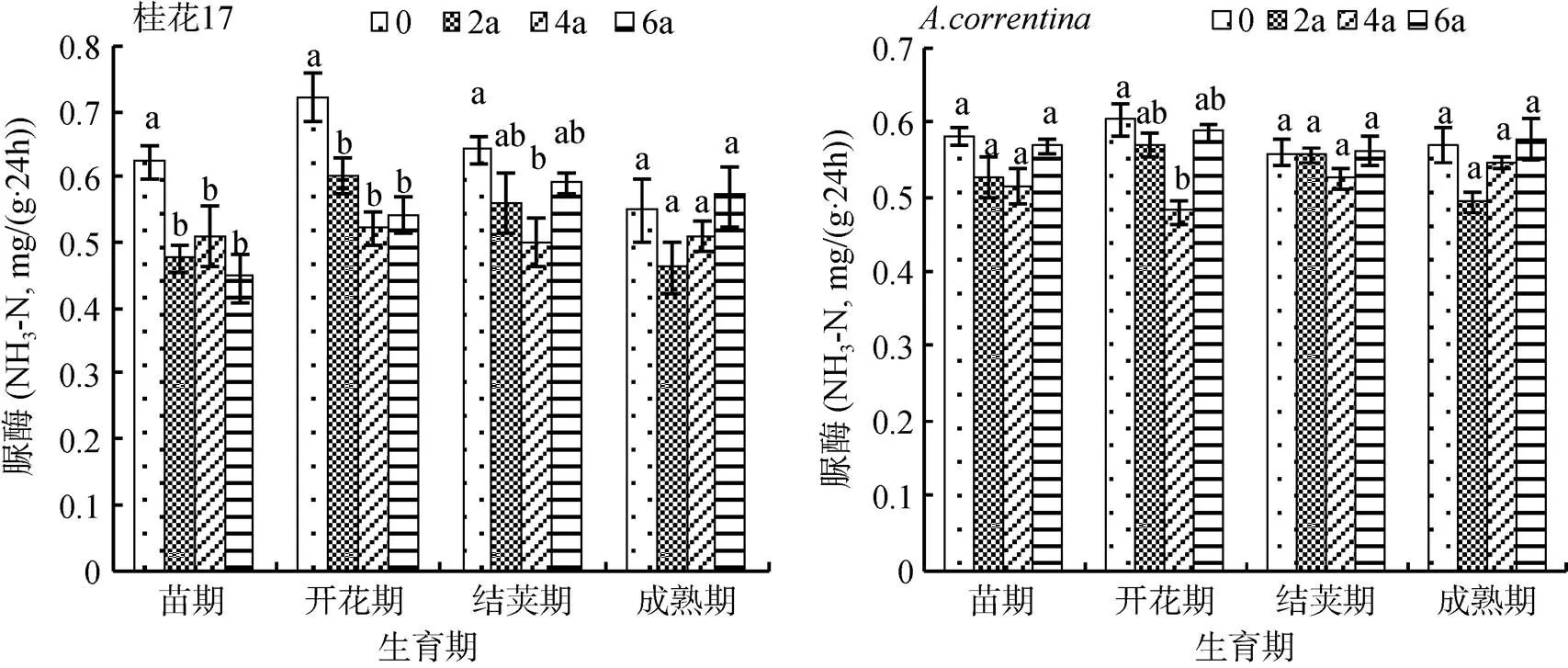
(图中不同小写字母表示同一花生品种同一生育期各处理间差异在P<0.05水平显著,下图同)
2.1.2 转化酶 由图2可知,桂花17花生各年限处理各生育期的土壤转化酶活性变化总体趋势为:CK>连作2 a>连作4 a>连作6 a。与CK相比,桂花17的土壤转化酶活性随着连作年限的增加而逐渐降低,以开花期和成熟期土壤转化酶活性下降最大。在开花期和成熟期,连作2 a处理分别比CK降低了15.9% 和24.6%,连作4 a处理分别比CK降低30.2% 和41.2%,连作6 a处理分别比CK降低11.6% 和39.5% (图2)。
而各生育期的土壤转化酶活性除了连作2 a处理均低于CK外,其他2个连作年限处理总体表现无一致性规律,总体表现为:连作2 a处理的土壤转化酶活性降低,连作4 a处理又增加,连作6 a处理降低。连作2 a处理与CK相比,苗期的土壤转化酶活性下降5.5%,开花期下降15.3%,结荚期和成熟期分别下降14.7% 和18.1%。
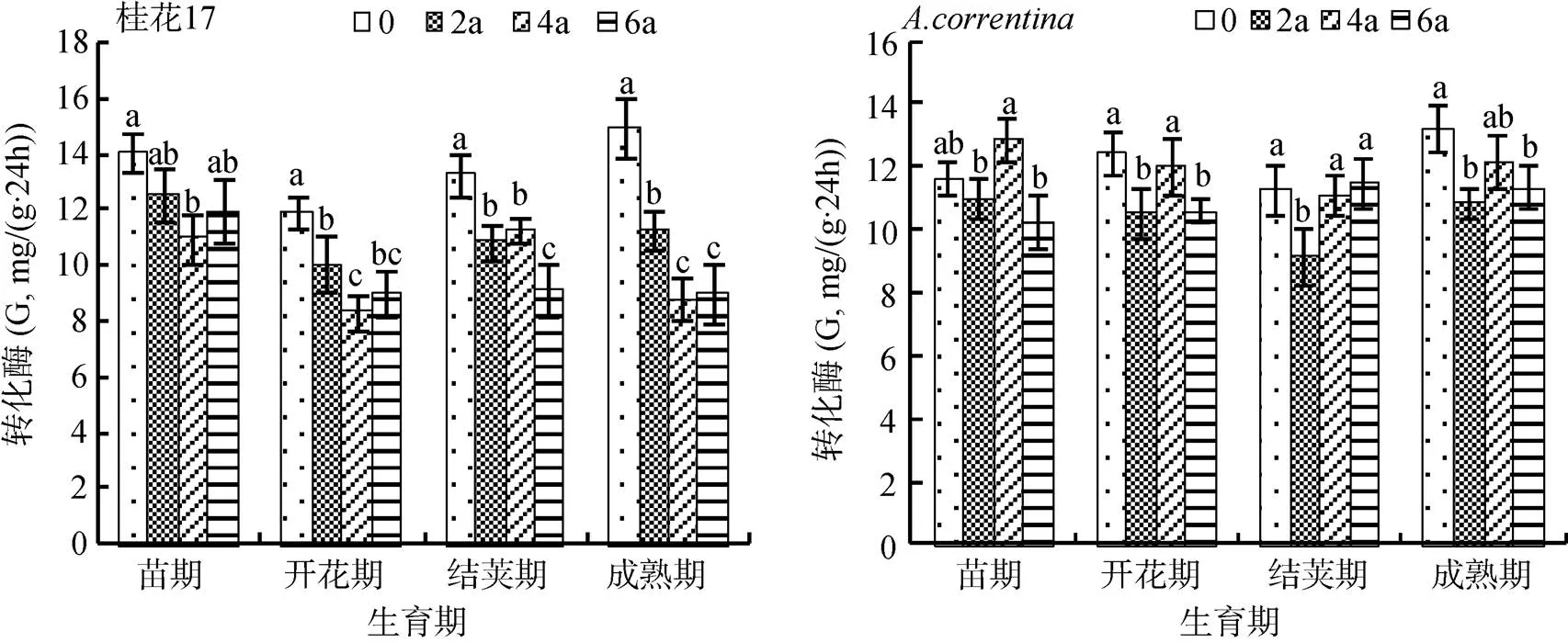
图2 花生连作对土壤转化酶活性的影响
2.1.3 酸性磷酸酶 由图3可知,桂花17各连作年限处理与CK相比,连作降低了桂花17各生育期的土壤酸性磷酸酶活性,各生育期总体上表现为:CK的土壤酸性磷酸酶活性最高,在连作2、4、6 a处理土壤酸性磷酸酶活性呈下降趋势,与土壤脲酶活性的变化趋势相似。在成熟期,各连作处理土壤酸性磷酸酶活性显著低于CK,连作2 a处理比CK下降22.2%,连作4 a处理下降51.9%,连作6 a处理下降44.3%(图3)。
与CK相比,各连作年限处理除了成熟期的土壤酸性磷酸酶活性低于CK外;其他3个生育期各连作年限处理的酸性磷酸酶活性或升高或降低,而且酶活性上升或下降幅度小,各处理间的差异大多不显著,表现无一致性规律。在成熟期,与CK相比,连作2 a处理土壤酸性磷酸酶活性比CK下降26.9%,连作4 a处理下降23.1%,连作6 a处理下降30.8%。
2.1.4 过氧化氢酶 由图4可知,与CK相比,两个花生品种在不同的连作年限间的土壤过氧化氢酶活性的变化并未随连作年限的增加表现出逐渐增加或降低的一致性规律。与CK相比,桂花17花生各生育期土壤过氧化氢酶活性总体表现为:连作2、4 a处理土壤过氧化氢酶活性先下降后上升,连作6 a处理土壤过氧化氢酶活性总体呈下降趋势。而花生各生育期总体表现为,连作2 a处理土壤过氧化氢酶活性上升到最高,连作4 a处理下降,连作6 a处理低于CK。
2.2 连作对花生成熟期土壤养分含量的影响
与CK相比,在花生成熟期桂花17各连作年限处理的土壤有效养分含量均降低,差异均达显著水平(表2)。与桂花17一致,不同连作年限土壤有效养分含量表现出降低趋势,其中连作6 a处理与CK的差异均达到显著水平(<0.05),但处理间变幅比桂花17小。表明由于花生连作造成了土壤养分含量的下降。
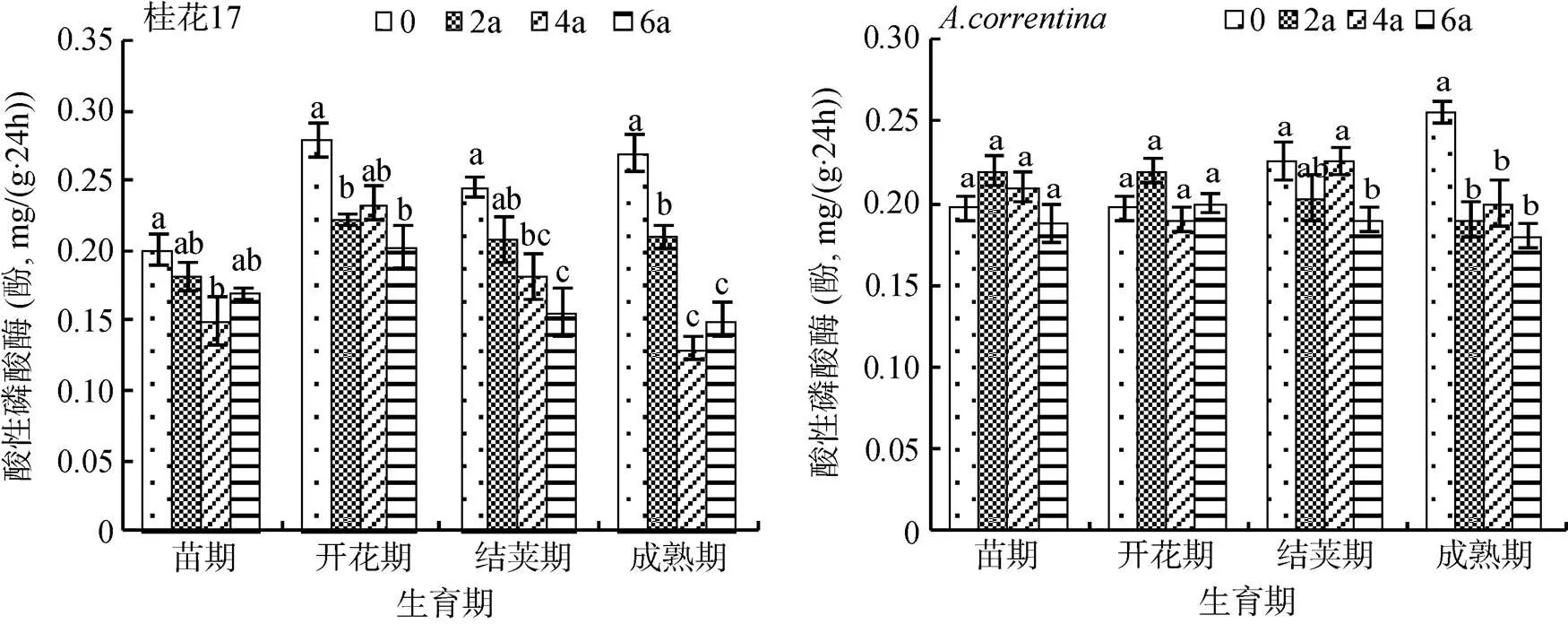
图3 花生连作对土壤酸性磷酸酶活性的影响
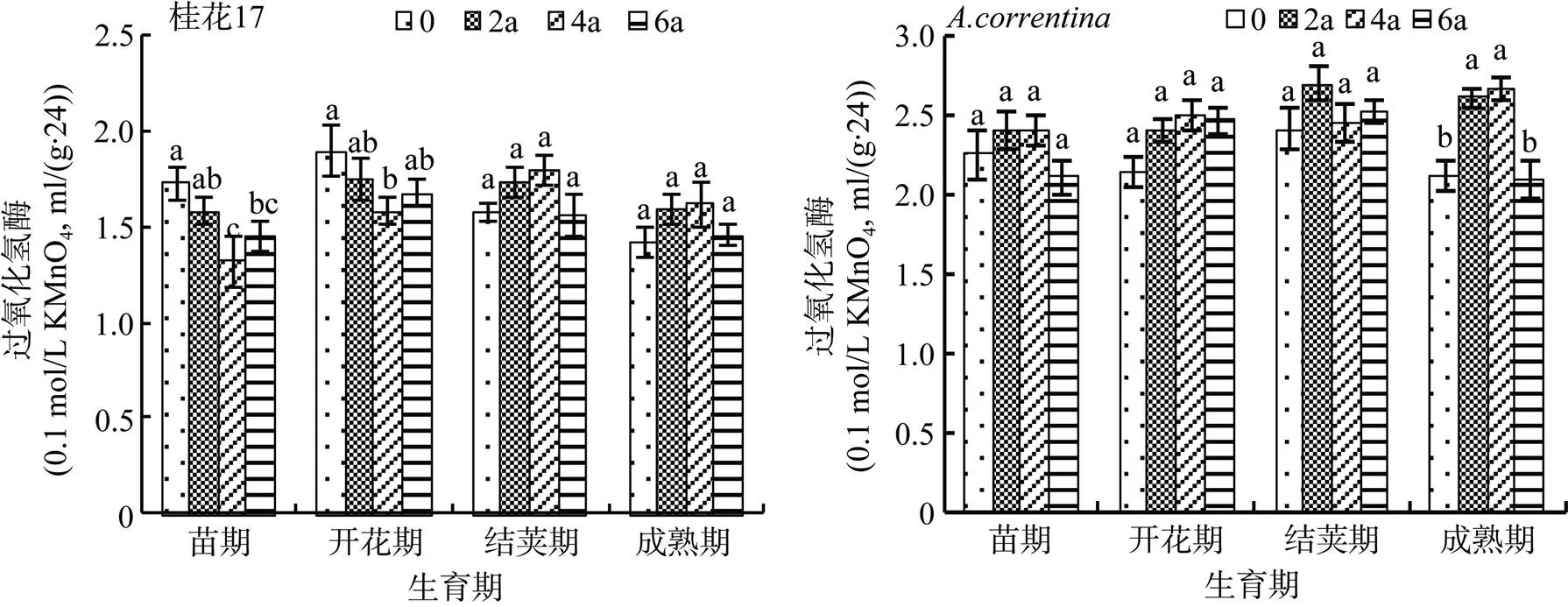
图4 花生连作对土壤过氧化氢酶活性的影响
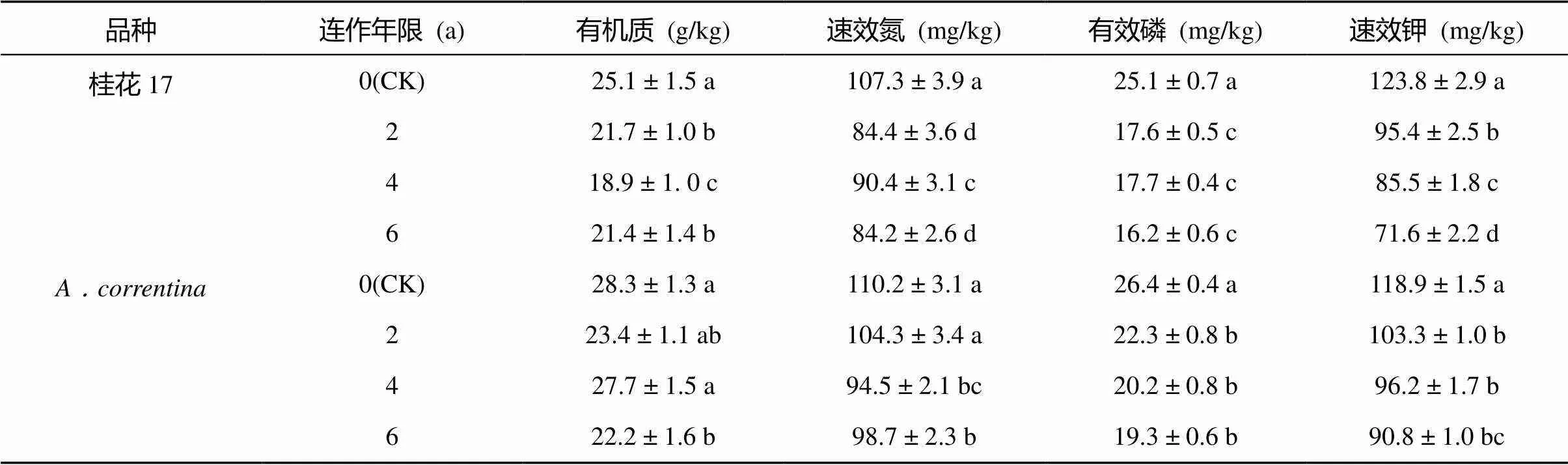
表2 花生连作对土壤养分含量的影响
注:同列数值后小写字母不同表示同一花生品种下各处理间差异在<0.05水平显著,下同。
2.3 土壤酶与土壤养分的相关关系
由表3可知,在花生成熟期两个花生品种土壤脲酶、转化酶和酸性磷酸酶活性与土壤养分指标间均存在显著相关关系,表明以上土壤酶活性可以作为衡量微生物活性和土壤肥力的指标,已有的研究也得出相似的结果[14-16]。桂花17除了土壤过氧化氢酶活性与土壤有机质含量不存在显著的相关关系外,其他土壤酶活性与各土壤养分含量间均存在显著(<0.05)和极显著(<0.01)正相关关系。而的土壤过氧化氢酶活性与各土壤养分含量指标均不存在显著的相关关系,的土壤脲酶和酸性磷酸酶活性与各土壤养分含量指标呈正相关关系,达显著水平(<0.05)。的土壤转化酶活性与土壤有机质含量呈显著正相关(<0.05)。
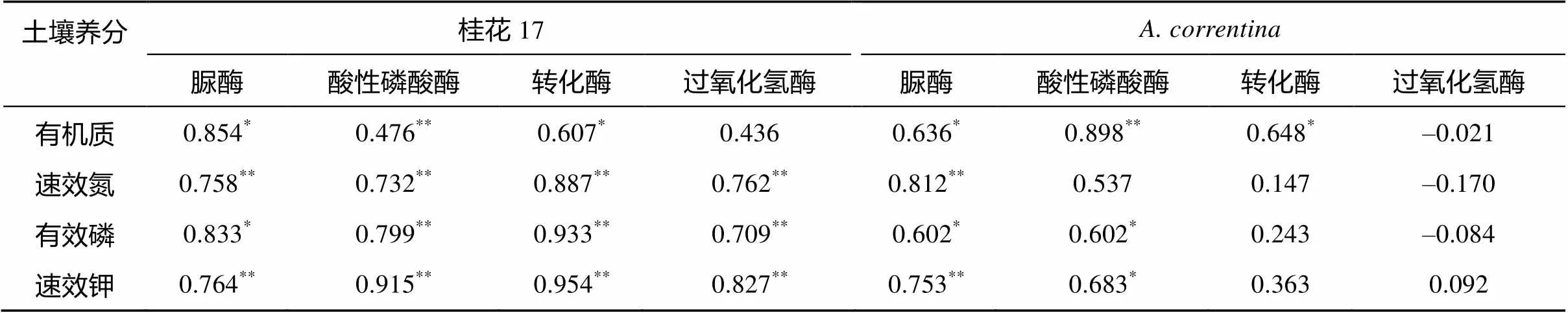
表3 花生土壤酶与土壤养分的相关关系
注:*表示在<0.05 水平显著相关(双侧);**表示在<0.01 水平显著相关(双侧)。
2.4 花生连作对干物质产量的影响
本研究以花生植株总干物质量作为产量考察指标,考察桂花17和的耐连作能力。结果表明(表4),各连作年限处理桂花17生长速度均比快速,单位面积生长量大。从本试验结果看,桂花17的CK和连作2 a处理的干物质产量无显著差异,但均显著高于连作4 a和连作6 a处理。随着连作年限增加降低了桂花17干物质产量,以连作6 a处理下降幅度最大,在苗期、开花期、结荚期、成熟期连作6 a处理比CK分别下降1.8%、14.2%、17.1% 和15.2%。干物质产量随连作年限增加呈下降趋势,但处理间差异均未达显著水平。与CK相比,连作6 a处理的干物质产量在苗期、开花期、结荚期、成熟期较CK仅分别下降2.2%、2.9%、–1.2% 和4.9%。分析表明,耐连作能力比桂花17强。
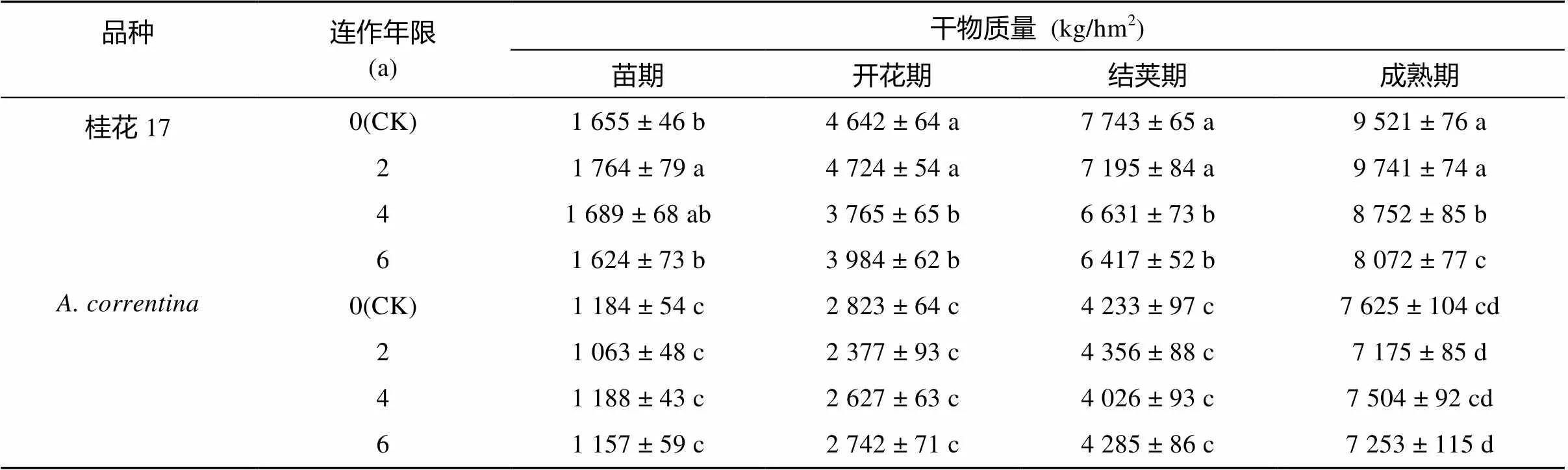
表4 连作对A. correntina和桂花17干物质积累的影响
3 讨论
3.1 连作对花生野生种和栽培种土壤酶活性的影响
关于土壤酶与连作障碍的研究,已经在大豆、棉花、烟草、草莓和黄瓜等作物进行了大量研究[17-20],但已有研究结果并不一致。连作对土壤酶活性的影响,因作物种类、栽培方式、土壤养分状况和土壤理化性状不同而表现出不同的变化规律。从本试验结果看,连作对和桂花17土壤酶活性影响不一致。花生各生育期土壤脲酶、转化酶、酸性磷酸酶活性总体表现无一致性上升或下降的规律,处理间的变幅比桂花17小。桂花17土壤脲酶、转化酶和酸性磷酸酶活性在各生育时期变化趋势基本一致,酶活性的变化总体上随连作年限的增加而表现出逐渐降低的趋势。说明连作降低了桂花17土壤的生物活性,不利于土壤中氮素和有机磷的水解矿化作用,从而降低土壤有效氮和磷的供应能力以及土壤中易溶性营养物质释放,影响了土壤的供肥能力。桂花17土壤酶活性的降低其原因可能是由于化感物质及致病性真菌的累积所致。孙秀山等[21]研究表明花生连作土壤中碱性磷酸酶、蔗糖酶和脲酶活性逐年降低,与本研究结果相似,也与赵萌等[22]有关连作土壤脲酶以及张淑香等[23]有关土壤转化酶的研究结果相似。黄玉茜等[24]研究表明随着花生连作年限的增加,在整个生育期内,转化酶活性在不同生育时期内变化规律不一致,土壤酸性和中性磷酸酶活性均呈现先升高后下降的趋势,与本研究结果不尽相同。顾美英等[25]研究表明连作土壤酶活性呈下降(过氧化氢酶和磷酸酶)或先升高后下降(脲酶和蔗糖酶)趋势,但随着连作年限的延长,这 4 种土壤酶活性均表现出增高趋势。本研究两个花生品种的土壤过氧化氢酶各连作处理间酶活性变化无规律性,与黄玉茜等[24]的研究结果相符。
在连作条件下可能通过影响土壤酶活性来调节其生态适应性,其原因一方面可能是根系分泌物差异造成。新近的研究表明,鉴定出的根系水培分泌物与花生栽培种桂花22存在差异,苯甲醛、对乙基苯甲酸和阿魏酸对的化感抑制作用也明显低于桂花22[26]。另一方面也可能是根际微生物活动及微生物区系差异造成。研究表明,与正茬相比,随着连作年限的增加,土壤细菌、放线菌数量和土壤有效养分含量相比桂花17下降幅度小,根际土壤的真菌数量相比桂花17增加幅度小[27]。
3.2 连作对花生野生种和栽培种土壤养分的影响
连作障碍普遍造成作物产量下降和土壤养分耗竭。研究表明连作造成花生栽培种植株对土壤养分选择性吸收,导致土壤养分不平衡,造成花生植株营养吸收障碍[24,28-29]。本研究表明两个花生品种不同连作年限土壤有效养分含量均降低,与前人的研究结果一致。相比桂花17,在连作条件下土壤有效养分含量下降幅度小,土壤养分吸收利用差异可能是在连作条件下相比桂花22有较好的耐受性的原因之一。研究也表明,相比花生栽培种,对磷、钾的敏感性更弱。的光合生产受磷、钾营养水平的影响相对较小,磷、钾素利用效率高于栽培种桂花22[27,30]。在改良花生栽培种营养利用效率方面有良好前景。
3.3 连作对花生野生种和栽培种植株产量的影响
本研究表明两个花生品种不同连作年限植株生物产量均降低,是造成花生连作障碍的主要原因之一。这与张艳君等[2]连作对花生生长发育有明显的抑制作用的研究结果相似。植株生产量差异可反映作物不同品种耐连作能力的差异。相比桂花17不同连作年限土壤有效养分含量和干物质产量相比下降幅度小,这可能是在连作条件下相比桂花17有较好的耐受性的原因之一。
由于目前关于土壤化感物质和土壤微生物的分离鉴定一直是个难点,分离出的数量和种类有限,研究结果还存在局限性,如何将其与土壤酶、土壤养分状况等指标结合研究连作对土壤微环境的影响,以全面探明连作条件下作物的土壤生理生态变化的机理机制是进一步研究的方向。另一方面还需通过考察其他与连作相关的生理生态及形态指标,如根系分泌物、植株养分吸收利用差异等分析,研究花生野生品种在连作条件下的形态生理机制及分子机理,为其进一步地有效利用于耐连作研究提供基础。
4 结论
和桂花17随着连作年限的增加,土壤有效养分含量和植株干物质产量均降低,相比桂花17下降幅度小。桂花17各连作年限处理的脲酶、转化酶、酸性磷酸酶活性随连作年限的增加有下降的趋势,处理间变幅大,而处理间变幅小。相关性分析显示两个花生品种的脲酶、转化酶和酸性磷酸酶活性与土壤主要养分间存在显著正相关,表明以上土壤酶活性在一定程度上可以作为衡量微生物活性和土壤肥力的指标。综上所述,在连作条件下,的土壤理化和生物学性状相比桂花17的变化幅度更小,具有更强的生理生态适应性。
[1] 王月, 刘兴斌, 韩晓日, 等. 不同施肥处理对连作花生土壤微生物量和酶活性的影响[J]. 沈阳农业大学学报, 2016, 47(5): 553–558
[2] 张艳君, 郭丽华, 于涛, 等. 花生连作对植株生长发育及主要农艺生理指标的影响[J]. 辽宁农业科学, 2015(6): 17–20
[3] 康亚龙, 景峰, 孙文庆, 等. 加工番茄连作对土壤理化性状及微生物量的影响[J]. 土壤学报, 2016, 53(2): 533–542
[4] 李鑫, 张秀丽, 孙冰玉, 等. 烤烟连作对耕层土壤酶活性及微生物区系的影响[J]. 土壤, 2012, 44(3): 456–460
[5] 冯国忠, 王寅, 焉莉, 等. 土壤类型和施氮量对连作春玉米产量及氮素平衡的影响[J]. 土壤学报, 2017, 54(2): 444–455
[6] 徐雪风, 李朝周, 张俊莲. 轮作油葵对马铃薯生长发育及抗性生理指标的影响[J]. 土壤, 2017, 49(1): 83–89
[7] Halward T M, Stalker H T, Larue E A, et al. Genetic variation detectable with molecular markers among unadapted germplasm resources of cultivated peanut and related wild species[J]. Genome, 1991, 34(6): 1013–1020
[8] 李忠, 江立庚, 唐荣华, 等. 花生栽野杂交后代抗病基因类似物的克隆与分析[J]. 西北农林科技大学学报: 自然科学版, 2014, 42(11): 32–38
[9] 唐荣华, 周汉群. 花生属栽野杂种后代抗青枯病研究[J]. 中国油料作物学报, 2000, 22(3): 61–65
[10] 张新友. 挖掘利用近缘野生种质加强花生种质创新[J]. 作物杂志, 2012(6): 6–7
[11] 周蓉. 花生(野生种)叶部病害抗病性鉴定[J]. 花生科技, 1990(2): 33–34
[12] 关松荫. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986
[13] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000: 147–149
[14] Kromer S, Green D M. Acid and alkaline phosphatase dynamics and their relationship to soil microlimate in asemiarid woodland[J]. Soil Biology Biochemistry, 2000, 32: 179–188
[15] Allison V J, Condron L M, Peltzer D A, et al. Changes in enzyme activities and soil microbial community compositional on gcarbon and nutrient gradients at the Franz Josefchrono sequence, New Zealand[J]. Soil Biology and Biochemistry, 2007, 39: 1770–1781
[16] 杨媛媛, 陈奇伯, 黎建强, 等. 滇中高原华山松植物多样性与土壤生物肥力特征[J]. 土壤, 2017, 49(1): 90–96
[17] 谷岩, 邱强, 王振民, 等. 连作大豆根际微生物群落结构及土壤酶活性[J]. 中国农业科学, 2012, 45(19): 3955–3964
[18] 李锐, 刘瑜, 褚贵新, 等. 棉花连作对北疆土壤酶活性、致病菌及拮抗菌多样性的影响[J]. 中国生态农业学报, 2015, 23(4): 432–440
[19] 尤垂淮, 曾文龙, 陈冬梅, 等. 不同养地方式对连作烤烟根际土壤微生物功能多样性的影响[J]. 中国烟草学报, 2015, 21(2): 68–74
[20] 田给林, 严婷婷, 毕艳孟, 等. 草莓连作土壤灭菌与施用有机肥对根际土壤酚酸及土壤酶活性的影响[J]. 园艺学报, 2015, 42(10): 2039–2048
[21] 孙秀山, 封海胜, 万书波, 等. 连作花生田主要微生物类群与土壤酶活性变化及其交互作用[J]. 作物学报,2001,27(5):617–620
[22] 赵萌, 李敏, 王淼焱, 等. 西瓜连作对土壤主要微生物类群和土壤酶活性的影响[J]. 微生物学通报, 2008, 35(8): 1251–1254
[23] 张淑香, 高子勤, 刘海玲. 连作障碍与根际微生态研究Ⅲ.土壤酚酸物质及其生物学效应[J]. 应用生态学报, 2000, 11(5): 741–744
[24] 黄玉茜, 韩立思, 韩梅, 等. 花生连作对土壤酶活性的影响[J]. 中国油料作物学报, 2012, 34(1): 96–100
[25] 顾美英, 徐万里, 茆军, 等. 新疆绿洲农田不同连作年限棉花根际土壤微生物群落多样性[J]. 生态学报, 2012, 32(10): 3031–3040
[26] 李忠, 江立庚, 唐荣华, 等. 花生根系分泌物的鉴定和化感作用研究[J]. 华南农业大学学报, 2015, 36(5): 48–53
[27] 李忠. 花生野生种耐连作生理基础研究[D]. 南宁: 广西大学, 2016
[28] 万书波, 王才斌, 户俊玲, 等. 连作花生的生育特性研究[J]. 山东农业科学, 2007(2): 32–36
[29] 封海胜, 张思芬,万书波, 等. 连作花生土壤养分变化及对施肥反应[J]. 中国油料, 1993(2): 53–57
[30] 李忠, 江立庚, 唐荣华, 等. 施钾量对花生光合特性及干物质积累的影响[J]. 贵州农业科学, 2016, 44(2): 52–55
Effects of Long-Term Continuous Peanut Cropping on Dry Matter Weight of Different Peanut Varieties, Soil Nutrient Contents and Enzyme Activities
LI Zhong1,2, JIANG Ligeng2, TANG Ronghua1,3, GUO Wenfeng1
(1 Guangxi Crop Genetic Improvement and Biotechnology Lab, Nanning 530007, China; 2 Agricultural College, Guangxi University, Nanning 530007, China; 3 Cash Crops Research Institute, Guangxi Academy of Agricultural Sciences, Nanning 530004, China)
A pot experiment was conducted to study the effects of continuous peanut cropping (0, 2, 4 and 6 years) on rhizosphere soil enzyme activities, soil nutrient contents and dry matter accumulation of cultivar Guihua17 and wild variety. The results showed that with the continuous cropping year increase, compared with the control (no continuous cropping), soil available nutrient contents and dry matter accumulation were decreased, the decline inwas lower than Guihua 17, the activities of soil urease, invertase and acid phosphatase of Guihua 17 decreased, the activities of soil urease, invertase and acid phosphatase ofat various stages changed irregularly, and little difference occurred between different treatments. The activites of soil hydrogen peroxide enzymes of the two varieties also changed irregularly. Comparatively, the changes in soil physical and chemical properties, biological properties of wild specieswas smaller than cultivars variety Guihua 17.
Peanut; Soil enzyme activity; Soil nutrients; Dry matter weight
广西青年基金项目(桂科青0991016)、现代农业产业技术体系建设专项资金(CARS-14-华南区栽培岗位)和广西重点实验室建设项目(12-071-09)资助。
李忠(1979—),男,广西百色人,博士,副研究员,主要从事作物栽培与遗传育种。E-mail:gxlizhong@126.com
10.13758/j.cnki.tr.2018.03.008
S565.2
A
猜你喜欢
东南大学学报(自然科学版)(2022年4期)2022-08-12
作物学报(2022年3期)2022-01-22
现代畜牧科技(2021年4期)2021-07-21
腐植酸(2020年1期)2020-11-29
中国现代药物应用(2020年10期)2020-06-18
当代工人(2019年22期)2019-12-20
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
安徽科技学院学报(2018年3期)2018-09-18
分析化学(2018年7期)2018-09-17
中成药(2017年6期)2017-06-13
