
山西历山山地草甸群落及种群格局
2018-07-20 12:14:28吴萍萍
东北林业大学学报 2018年7期
吴萍萍
(山西省林业调查规划院,太原,030012)
群落分布格局是由群落内各个种格局斑块相互交错重叠构成[1-2]。种群格局的形成既与物种自身的生物学特性有关,还与种间关系、气候因子、地形因子和土壤因子等环境因子以及人类活动密切相关[1]。目前,植物群落和种群的空间格局分析已成为研究植物动态、种间关系以及物种与环境关系的一种重要手段。关于格局分析的方法已有很多,并在种群和群落格局研究中得到广泛应用[3-11]。种群斑块的形成、动态都会对整个群落产生不同程度的影响,优势种的格局变化,很大程度上决定了群落的结构和动态[2]。因此,研究群落及其优势种的分布格局对于了解群落的组成、结构以及演替等都有重要意义。张金屯[12]和李素清等[13]应用除趋势对应分析(DCA)与双项轨迹方差法(TTLQV)相结合的方法,分别对芦芽山和云顶山亚高山草甸群落及其优势种的格局进行分析,结果表明DCA和TTLQV相结合的方法可以客观地反映群落及其优势种的空间分布。群落分布格局与其优势种分布格局关系密切,其分布格局不仅与物种的生物学特性有关,还受种间关系、环境因子及人类活动的影响。
山西历山舜王坪是华北典型的山地草甸,具有明显区系组成和丰富植物多样性,然而长期无序的旅游造成山地草甸严重的生态环境问题,包括植被盖度的下降、生物多样性减少以及严重的水土流失等[14-15]。车前(PlantagoasiaticaL.)作为一种典型的伴人植物,车前的分布格局和种群数量、动态可以指示人类活动对生态系统的干扰情况[16]。因此,研究车前种群的分布格局变化,对于揭示人类活动对群落和种群的分布格局动态具有一定的科学意义。因此,本文采用DCA与TTLQV相结合的方法,研究群落和优势种群格局沿海拔梯度的变化规律,同时选取典型的伴人植物车前作为研究对象,通过TTLQV方法分析其格局变化,分析旅游活动等干扰对植物种群分布格局的影响,进而揭示人类活动与群落和种群分布格局的动态关系。
1 研究区概况
历山国家自然保护区位于山西中条山东段,地理位置:111°51′~112°30′E,35°16′~35°27′N。区域地貌属于侵蚀构造中山,海拔650~2 358 m,相对高差达1 708 m。该区属暖温带大陆性季风气候,年均气温13.1 ℃,≥10 ℃以上有效积温4 281 ℃,年降水量600~800 mm。土壤类型主要包括褐土、山地褐土、棕色森林土和山地草甸土[14-15]。森林群落主要包括栓皮栎(QuercusvariabilisBI.)林、辽东栎(Quercuswutaishanica)林、桦木(Betulaspp.)林、山杨(Populusdavidiana)林和油松(Pinustabuliformis)林等。灌丛群落主要包括荆条(VitexnegundoL.)灌丛、黄刺玫(RosaxanthinaLindl)灌丛、虎榛子(OstryopsisdavidianaDecaisne)灌丛和连翘(Forsythiasuspensa)灌丛等。草本群落主要包括白羊草(Bothriochloaischaemum(L.) Keng.)草丛、蒿类(Artemisiaspp.)草丛和薹草(Carexspp.)草甸等[17]。
舜王坪山地草甸位于历山主峰舜王坪,海拔2 358 m。地理位置:111°57′~111°58′30″E,35°23′45″~35°25′25″N。群落类型主要有薹草草甸、紫羊茅(Festucarubra)草甸和五花草甸(Festucarubra+Carexspp.+Sanguisorbaofficinalis)等,优势植物主要包括紫羊茅(Festucarubra)、草地早熟禾(Poapratensis)、早春薹草(Carexsubpediformis)和地榆(Sanguisorbaofficinalis)等[18]。
2 研究方法
2.1 样带设置
2004年8月,采用样带法分别在历山舜王坪海拔2 100、2 150、2 200、2 250、2 300 m的山地草甸各设置1条样带,共5条样带(样带1、样带2、样带3、样带4、样带5)。每条样带长12.8 m,由128个10 cm×10 cm连续小样方组成,记录每个小样方中所有物种的种名、盖度和高度等。
2.2 分析方法
运用TTLQV方法分别对5条样带的优势种和车前进行格局分析,区组方差的公式如下[19-20]:
式中:V(b)为区组方差,b为区组,j为起始样方的序号,xi为样方i中物种的盖度,n为样方数。
区组大小表示所含样方的个数,最大的区组为总样方个数的一半,即64(128/2)。首先分别计算物种在每个区组下所对应的区组方差,然后以区组大小为横坐标,区组方差(V(b))为纵坐标绘制优势种格局分布图,V(b)峰值所对应的区组大小即为物种在该尺度下的格局规模。本文最大的格局规模为总样带长的一半,即640 cm,格局规模大小按照等间距标准进行划分[20],即10~210 cm为小格局、210~400 cm为中格局、410~640 cm为大格局。
应用DCA分别对5条样带所有种进行排序,分别得到每条样带128个样方的DCA排序坐标值。以小样方为横坐标,分别以DCA第1、第2排序轴为纵坐标,绘制排序值变化图,得到群落格局分析图,对比它们之间的结果,用以描述群落的格局分析。群落分类采用双向指示种分析法(TWINSPAN)。
2.3 群落类型
依据《中国植被》的群落命名原则[21],采用优势种命名法,结合双向指示种方法分类结果,样带所在的草甸群落分别命名为:紫羊茅+草地早熟禾草甸(Ass.Festucarubra+Poapratensis)、早春薹草+紫羊茅草甸(Ass.Carexsubpediformis+Festucarubra)、草地早熟禾+早春薹草草甸(Ass.Poapratensis+Carexsubpediformis)、等穗薹草+草地早熟禾草甸(Ass.Carexleucochlora+Poapratensis)、龙胆+等穗薹草甸(Ass.Gentianascabra+Carexleucochlora)。群落中的建群种、优势种和伴生种的确定依据TWINSPAN分类结果和物种出现的总样方数以及重要值进行划分。
3 结果与分析
3.1 优势种的格局
由图1可知,样带1(紫羊茅+草地早熟禾草甸类型)共建种紫羊茅和草地早熟禾分别在区组60和55处有明显的峰值,表明紫羊茅和草地早熟禾具有明显的大斑块格局,规模大小分别为600和550 cm,而在区组6和7处的峰值不明显,表现出规模大小为60和70 cm的小斑块。优势种车前和蒲公英的斑块峰值均不明显,大斑块分别出现在区组62和58处,规模分别为620和580 cm,小斑块分别出现在区组8和5处,规模大小分别为80和50 cm;建群种和优势种的中斑块均未表现出来。

a.紫羊茅(Festuca rubra);b.草地早熟禾(Poa pratensis);c.车前(Plantago asiatica);d.蒲公英(Taraxacum mongolicum)。
由图2可知,样带2(早春薹草+紫羊茅草甸)共建种早春薹草和紫羊茅仅呈现出小斑块,分别在区组6和4处出现峰值,规模大小分别为60和40 cm,无大斑块和中斑块。优势种地榆的小斑块规模为50 cm,在区组19处,有一个近似中斑块的峰值,斑块大小为190 cm,大斑块为570 cm;优势种东方草莓在区组2和35处有峰值,表明该种的小斑块和中斑块的规模分别为20和350 cm。
由图3可知,样带3(草地早熟禾+早春薹草草甸类型)共建种草地早熟禾有两个小斑块,分别出现在区组2和11,规模大小分别为20和110 cm,大斑块出现在区组59处,规模大小为590 cm;共建种早春薹草在区组2和62处的峰值不明显,表明该种斑块格局不明显。优势种蒲公英在区组2和15处表现出小斑块格局,格局规模为20和150 cm,中斑块格局不明显,大斑块出现在区组63处,规模大小为630 cm;优势种东方草莓的大斑块规模为620 cm,中斑块规模为260 cm,并未出现小斑块格局。
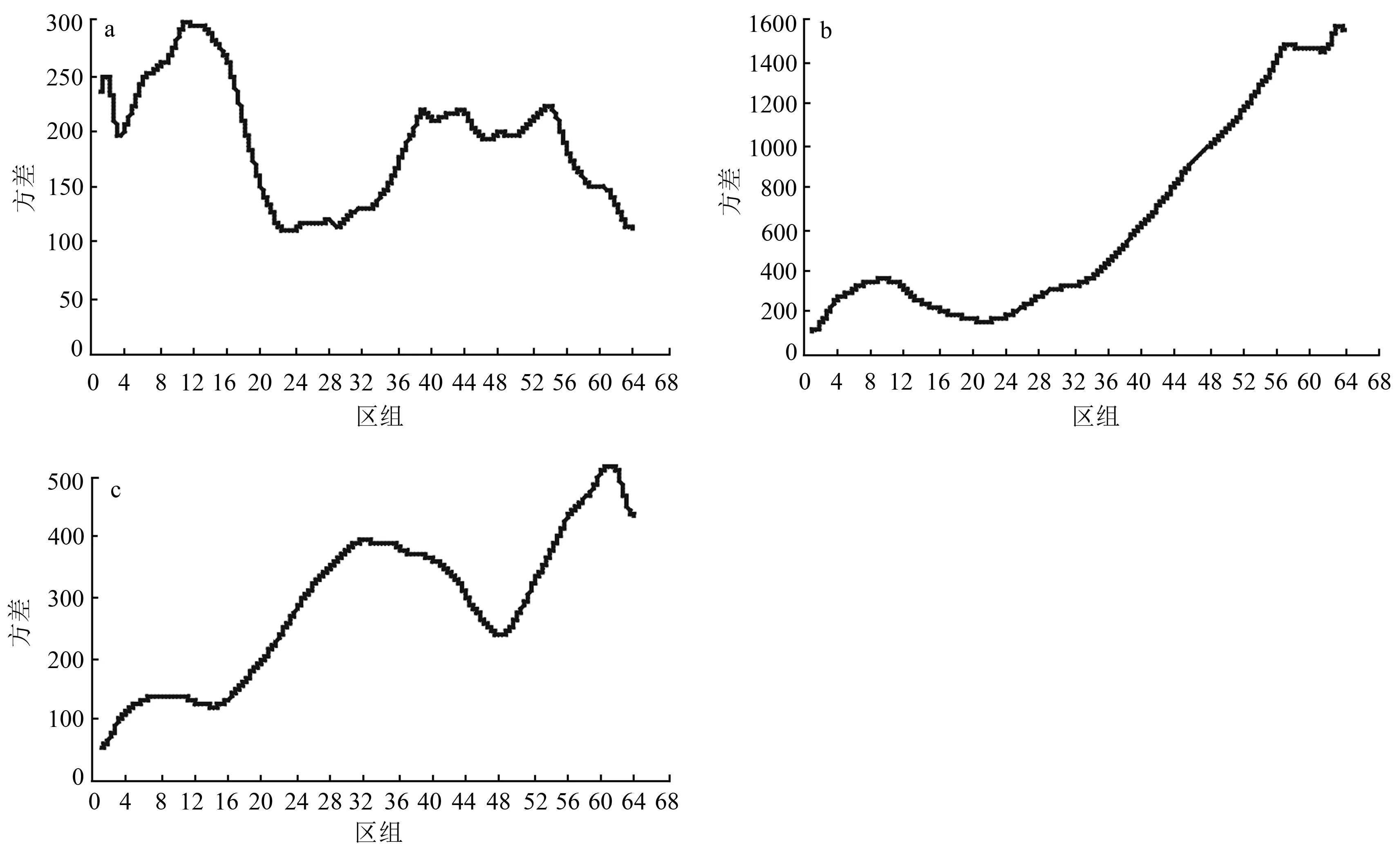
由图4可知,样带4(等穗薹草+草地早熟禾草甸)共建种等穗薹草的小斑块分别出现在区组2和11,规模大小分别为20和110 cm,中斑块和大斑块分别出现在区组39和54处,规模大小分别为390和540 cm;共建种草地早熟禾的大斑块规模为570 cm,小斑块规模为90 cm。优势种东方草莓的大斑块规模为610 cm,中斑块规模为320 cm,小斑块规模为90 cm。
由图5可知,样带5(龙胆草甸)建群种龙胆在区组58处出现峰值,表明该种的大斑块规模为580 cm;优势种等穗薹草的大斑块规模为550 cm;这两个种均未出现中斑块和小斑块格局。
综合5条样带建群种与优势种的格局分布可知,历山山地草甸建群种的格局主要为大格局,而优势种的格局变化较多,这与种群的生物学特性、环境因子、群落内的种间关系等有关。如样带1的紫羊茅、样带2的早春薹草等建群种,一方面自身有较强的繁殖能力,另一方面对环境有较强的适应能力,可以优先占用资源。因此,在群落中分布广泛,占有主导地位,形成的格局规模主要为大格局。优势种或伴生种由于自身的生物学特点以及种间关系等因素,形成的格局往往出现在建群种斑块的间隙,格局规模主要表现为中、小格局,这种格局的镶嵌结构有利于群落内资源的充分利用和生态位互补,从而有助于群落格局和结构的稳定性。如样带2的共建种早春薹草的格局规模为大格局,而优势种地榆往往生长在早春薹草的格局间隙中,表现出小斑块规模(50 cm)。

a.早春薹草(Carex subpediformis);b.紫羊茅(Festuca rubra);c.地榆(Sanguisorba officinalis);d.东方草莓(Fragaria orientalis)。

a.草地早熟禾(Poa pratensis);b.早春薹草(Carex subpediformis);c.蒲公英(Taraxacum mongolicum);d.东方草莓(Fragaria orientalis)。
随着海拔梯度的变化,样带的物种组成、群落结构等方面在不同程度上会发生变化,群落与种群的格局也会随之改变。一方面,同一种在不同群落所处的地位和作用不同,物种的格局会发生改变。如等穗薹草在样带5中表现为大格局;而在样带4中,等穗薹草的格局多样,小斑块规模分别为20和110 cm,中斑块和大斑块规模分别为390和540 cm;另一方面,即使同一种在各群落中所处的地位相同,由于各群落的物种组成、种间关系等不同,物种形成的格局也会发生变化。如东方草莓在样带2中主要呈现“单峰”的中格局,在样带3中表现为大、中型的“双峰”格局,而在样带4中表现为大、中和小型的“三峰”格局。
3.2 群落的格局
由图6可知,根据DCA第1、第2排序轴得到的格局变化图,反映了5条样带沿海拔梯度群落格局的规模。综合DCA排序轴格局分布图可以看出,第1、第2排序轴的群落格局规模基本一致。
a.等穗薹草(Carex leucochlora);b.草地早熟禾(Poa pratensis);c.东方草莓(Fragaria orientalis)。

a.龙胆(Gentiana scabra);b.等穗薹草(Carex leucochlora)。
由图7可知,DCA排序轴格局分布图和TTLQV群落格局分布图,在反映群落格局变化方面有很好的一致性。样带1的群落主要呈现大斑块格局,规模大小为580 cm。样带2的群落格局主要为中格局,出现在区组35(350 cm),大斑块格局并未出现(图6)。样带3群落的小格局出现在区组5~10(50~100 cm),大格局出现在区组52~62(520~620 cm)。样带4群落形成的格局多样化,大、中和小格局均有出现。小格局发生在区组2(20 cm)和10(100 cm)处,中斑块为区组26(260 cm)或41区组(410 cm),大格局规模为610或630 cm。样带5群落的小格局出现在区组12(120 cm),大格局出现在区组53(530 cm)。
综合群落与种群的格局分布可以看出,群落格局沿海拔的变化与建群种的格局变化基本一致。这一结果表明群落的格局在很大程度上取决于群落的建群种及其优势种。群落格局实际上是群落内各物种格局的镶嵌结构[2,6],各物种的格局斑块相互交错重叠,共同构成了群落格局。
3.3 车前格局
由图8可知,垂直方向上,即从低海拔到高海拔(样带1到样带5),车前在样带中出现的频度及其盖度都有明显的减少;水平方向上,沿着各条样带,车前频度与盖度主要出现在前64个小样格。
在低海拔地区,人为干扰强度较大,样带1中,车前出现的频数最多(53次),盖度大多在10%以上,主要出现在前80的小样格中,表现出一种大尺度的“聚集”格局。样带1中车前的格局规模大小为620 cm。相对于样带1来说,样带2的人为干扰强度相对较弱,车前主要集中在某些小样格,表现出一种中小尺度的“聚集”格局。格局规模主要为中小格局(区组19),规模大小为190 cm。样带3的人为干扰活动相对较少,车前盖度多为5%;车前主要出现在前64的小样格中,格局规模表现为大格局(560 cm),表现出一种大尺度的“随机分布”格局。样带4和样带5由于海拔相对较高,人为干扰很少,车前的频度与盖度相对较低,分别形成的格局以大中尺度的随机分布(区组45和63)和中小尺度的随机分布(区组4和30)为主,格局类型变化多样。

a.第1条样带;b.第2条样带;c.第3条样带;d.第4条样带;e.第5条样带。

a.第1条样带;b.第2条样带;c.第3条样带;d.第4条样带;e.第5条样带。

a.第1条样带;b.第2条样带;c.第3条样带;d.第4条样带;e.第5条样带。
4 结论与讨论
历山山地草甸的格局变化分析表明:DCA排序与TTLQV相结合的方法可合理、客观地揭示历山山地草甸植物群落的格局特征,结果与DCA第1、第2排序轴反映的群落格局基本一致。张金屯[12]和李素清等[13]分别对芦芽山和云顶山的亚高山草甸的格局的研究结果类似。
沿海拔梯度,历山山地草甸群落的格局变化与建群种的格局变化基本一致。这表明群落的分布格局与建群种的格局密切相关。群落分布格局是群落内各种群格局分布的综合反映,是群落内各物种格局的镶嵌结构[2],其中,建群种在群落格局的形成过程中起着决定性作用。
随着海拔梯度的变化,样带的物种组成与群落结构等也会发生变化,进而导致群落和种群的格局发生改变。一方面,由于各群落的物种组成、种间关系等原因,即使同一物种在各群落中所处的地位相同,但形成的格局也会不同。如东方草莓在样带2、样带3和样带4均为优势种,但分布格局不尽相同。在样带2主要呈现“单峰”的中格局,在样带3表现为大、中型的“双峰”格局,而在样带4表现为大、中和小型的“三峰”格局。东方草莓在不同群落中分布格局不同表明种群的分布格局不仅与自身的生物学特性有关,同样受到海拔、土壤性质、人类活动以及群落内种间关系的影响。样带2处于低海拔区域,由于受到人为活动的干扰强度较大,导致一年生植物和伴人植物大量定居并迅速繁衍和生长,严重抑制了东方草莓的繁殖和克隆生长,导致其格局逐渐转化为离散化的中小斑块。相反,样带4由于人为干扰较少,东方草莓能够凭借较强的克隆繁殖能力占据有利空间,表现出较大的格局分布。另一方面,同一种在不同群落中扮演的角色、地位和作用不同,物种形成的格局也会有所不同。如等穗薹草在样带5的格局表现为大格局,而在样带4等穗薹草格局多样,小斑块规模分别为20和110 cm,中斑块和大斑块规模分别为430和540 cm。虽然样带4等穗薹草为群落建群种,但与样带5的等穗薹草相比,其大格局并不明显,这主要是因为这两个样带种间竞争和所处的演替阶段不同。样带4中草地早熟禾与等穗薹草处于相同的空间生态位上,与等穗薹草相比,草地早熟禾具有更强的竞争能力,可以更好地利用资源,占据有利的空间。随着草地早熟禾种群的扩大,等穗薹草逐渐会退居次要地位,格局规模将逐渐会转变成中小尺度的斑块。而样带5等穗薹草和龙胆由于空间生态位的差异,可以更好地相互依存,彼此都表现出大尺度的格局,这表明种间竞争以及在演替阶段所处的地位不同,种群所表现的格局也会有所不同。
山地草甸由于海拔相对较高,热量条件较差、生长季短,与低海拔地区草本植物群落相比,组成与结构相对简单,一旦遭到破坏,由于种质资源缺乏和生境恶化,短时间难以恢复,造成植被退化和生产力的迅速下降。多年来的过度放牧及近年来迅速发展的旅游业,对历山山地草甸植被和群落组成产生了明显的不良影响[14-15]。车前作为一种伴人植物,并非历山山地草甸的原生种,由于放牧或者人类旅游活动干扰,导致车前随人类或牲畜“登上”海拔2 000 m以上的历山山地草甸。车前在样带1出现频率最高,表现出一种大尺度的“聚集”格局,这主要是人为活动干扰严重所致。样带2的人为活动相对较轻,车前主要在某些小样格中集中分布,表现出一种中小斑块化的“聚集”格局。样带3的人为活动相对较少,表现出一种大尺度的“随机分布”格局。样带4和样带5由于海拔相对较高,人为干扰很少,车前的频度与盖度相对较低,分别形成的格局以大中尺度的“随机分布”和中小尺度的“随机分布”为主。从车前分布格局可以看出,其主要受人为活动强度与频度的影响。在干扰严重的低海拔区域(样带1和样地2),人类活动往往聚集在某一片区域,车前分布格局往往表现出集聚分布;而在干扰较弱的高海拔地区(样带4和样地5),车前分布格局随人类活动的随机性呈现随机分布。由此看以看出,人类活动会明显影响历山山地草甸格局类型的变化,主要表现在人类活动(包括旅游、放牧等)对群落的组成、种群数量和分布等方面,进而影响了群落的格局类型。因此,在山地草甸开发生态旅游的同时,尤其是对脆弱的生态环境,应当采取一定的保护措施,如规划旅游路线、修建游览栈道、科学制定生态旅游环境容量、加强生态环保意识,加大宣传力度等[18,22],最大限度地降低人类活动对山地草甸生态系统的不良影响,确保草甸生态系统的健康和可持续发展。
猜你喜欢
中国卫生统计(2022年4期)2022-10-12 01:55:12
——平衡不完全区组设计定量资料一元方差分析
四川精神卫生(2022年2期)2022-05-09 10:46:04
天津中医药大学学报(2020年1期)2020-03-03 14:00:42
青年生活(2019年32期)2019-09-10 04:25:38
小火炬·阅读作文(2019年4期)2019-08-06 04:33:28
畅谈(2018年24期)2018-02-02 02:31:16
民间文化论坛(2016年2期)2016-12-01 05:41:38
生态学报(2016年12期)2016-08-09 05:52:56
中国水土保持科学(2016年3期)2016-07-19 03:51:24
生态学报(2016年1期)2016-03-10 06:03:14
