
古尔班通古特沙漠南部植物多样性的区域差异
2016-08-09 05:52:56刘尊驰
生态学报 2016年12期
罗 宁,刘尊驰,于 航,刘 彤
石河子大学生命科学学院,石河子 832000
古尔班通古特沙漠南部植物多样性的区域差异
罗宁,刘尊驰,于航,刘彤*
石河子大学生命科学学院,石河子832000
摘要:植物多样性分布受环境影响,具有尺度依赖性,研究不同区域植物多样性的差异和分布规律有助于剖析群落多样性的整体形成和变化原因,进而为实施有效的保护和利用提供科学依据。以古尔班通古特沙漠植物多样性集中分布的南部为研究区,沿经向方向划分3条样带(绿洲边缘样带、克拉玛依至吉木萨尔的沙漠公路样带以及距离二者之间的沙漠腹地样带),同时根据沙垄高度、走向和生境差异将研究区划分为5个分区,对比研究各样带、各分区的α、β多样性特征及物种共性和差异性。结果表明:3条样带的α多样性变化为绿洲边缘样带>沙漠公路样带>沙漠腹地样带。随经度的增加,各样带的物种相似性和替代性特征表现相似的变化趋势。物种分析发现沙漠公路样带内短命植物分布较多,绿洲边缘样带以多年生草本和灌木群落为主,而沙漠腹地样带介于二者之间。各分区间的物种组成存在一定差异。沙漠西部、东部、莫索湾垦区附近、沙漠腹地等分区物种丰富度均较高,而古玛纳斯湖区的植物丰富度较低,与其余4个分区间的物种相似性小,相互替代性较强。总结认为古尔班通古特沙漠南部各分区间植物多样性受地史过程的影响较大,而样带间植物多样性受土壤质地、降水量等环境因素影响较大;沙漠东部似是整个南部沙漠的物种扩散中心。
关键词:沙漠;样带;分区;α多样性;β多样性;种库假说
在全球气候变化背景下,物种多样性的现状和变化是目前国内外生态学研究的热点之一。植物多样性分布具有尺度依赖性,在区域尺度下,物种多样性由于受到气候、干扰、生境类型等因素的影响,表现出不同的变化趋势[1- 4]。如今关于物种多样性的大尺度格局已提出了超过100种假说[5],例如 Rapoport 法则[6]、能量假说[7]、中间膨胀效应假说(Middle-Domain Effect, MDE)[8- 9]、种库假说[10]等,这些假说对物种产生不同分布格局的原因做出了巨大的贡献,但各假设得出的结论并不一致,也受到了较大的争议。区域生物多样性的形成与发展是由历史、地理、进化和生态过程等共同作用的结果。由于生境异质,区域植物多样性是由不同尺度的亚区域或局域多样性构成,而各尺度多样性的形成机制也各不同[11- 12],因此研究亚区域或局域的多样性特点对于全面了解区域生物多样性分布格局及其形成机制具有极大促进作用。
沙漠植物是沙漠生态环境保护的重点对象,植物多样性是沙漠生态系统稳定的基础。目前国外研究主要集中在分析沙漠地区的植物组成,植被层片功能以及区系特征,阐明地理、环境因子对沙漠区域植物分布格局的影响,剖析沙漠地区的多样性格局特点[13-16]。国内主要关注的是沙漠植物群落特征[17],群落物种组成[18],地理或环境因子等对植物群落格局的影响[19- 20]等方面,从不同区域角度分析群落整体植物多样性特征和原因的研究还较少。
古尔班通古特沙漠是世界温带具有代表性的沙漠之一,位于准噶尔盆地中央,面积4.88万km2,沙丘类型多样,地貌变化剧烈[21]。植物多样性较其他沙漠丰富,单种科、单种属多,区系优势现象明显[18],是我国极端环境植物种质资源库,但是近年来由于油田开采、道路兴建、放牧、地下水位下降等干扰,植被局部退化严重[22-23]。现有研究表明本沙漠植物分布呈现出明显的空间分异和多样性变化[24- 25]。但该沙漠面积较大,是由索布古尔布格莱沙漠、霍景涅里辛沙漠、德佐索腾艾里松沙漠、阔布北-阿克库姆沙漠4个小沙漠构成。由于海拔、距离绿洲和湖泊的远近差异,在土壤、地形、降水等存在明显的空间分异性,从不同区域上分析植物多样性特点和差异研究较少[26]。为此,本研究以古尔班通古特沙漠植物集中分布的南部为重点,将研究区自靠近绿洲到沙漠腹地,按经向方向划分3条样带;同时根据沙丘大小、走向、土壤类型、海拔、降水等将研究区划分为5个分区;对比不同样带、不同分区的植物多样性差异,总结沙漠植物多样性的变化规律,丰富古尔班通古特沙漠植物多样性研究,为合理保护该沙漠珍贵植物资源提供依据。
1材料与方法
1.1研究区概况
研究区位于准噶尔盆地南部,自西向东海拔从420—255—699 m呈中间凹形的特点。因受气流、盆地地形、水文和植被等因素的影响,沙漠地貌形态和植被类型出现明显的东西和南北方向上的分异[21,27],尤其沙漠东端高海拔地区降水达400—600 mm。整个研究区属典型的大陆性、干旱沙漠性气候。
1.23条样带的划分
根据多年对古尔班通古特沙漠全面调查的基础上,发现距离绿洲不同的远近上,植被类型和海拔等有较大差异。因而本研究根据距绿洲的远近,自绿洲向沙漠腹地沿经向上划分出3条样带(图1),即沙漠公路样带(或沙漠中线样带44°54′—45°30′ N);沙漠腹地与绿洲中间样带(简称沙漠腹地样带44°30′—45°18′ N),位于沙漠腹地深处,交通条件差,多样性调查困难,除油田开采影响外,放牧等人为干扰作用较弱;靠近绿洲的沙漠边缘样带(简称绿洲边缘样带44°23′—44°54′ N),处于绿洲-荒漠过渡带区域,植物多样性受到绿洲小气候的影响,水分充足,丰富度高。
1.35个分区的划分
根据古尔班通古特沙漠不同区域的地形、沙丘类型、海拔、降水量、植物分布特点将研究区划分成5个分区(图1)。分区一位于沙漠西部,与人工绿洲交错,受绿洲小气候的影响,其生存条件比较优越,植被生长较好。分区二位于莫索湾垦区附近,树枝状高大沙丘,短命植物多样性较为丰富[28]。分区三位于沙漠腹地,沙丘高大,新月型沙丘链与蜂窝状沙丘交错,由于生境、气候条件单一,受到人为破坏和干扰少,物种较为丰富。分区四位于沙漠东部的高海拔区(600 m以上),该区域降水较丰富,植物盖度大,且植被分布非常均匀。分区五位于古玛纳斯湖区,以平原为主,有2—3 m的低矮沙丘,受采油等人为破坏以及地下水位下降等影响[23],土壤次生盐渍化严重,且降水稀少,植被盖度低。

图1 研究区概况Fig.1 Survey of study area图中的3条粗线标记为3条样带
1.4样方设置和调查方法
根据临界抽样面积结果[29],以垂直于沙垄走向的方向设置100 m × 10 m(长×宽,10个10 m × 10 m的小样方)的样带,将调查的样方大小设定为1000 m2。在对研究区的全面踏查的基础上,对各样带和各分区内物种多样性变化大的地点布置样方,并由此产生相应的样方数量,所以从空间分布上看样点并不均匀,但通过此方法调查的结果基本反映了各分区的多样性特点。
各样带与各分区内的样方数分别为:沙漠公路样带有20个样方、沙漠腹地样带有16个样方、绿洲边缘样带有14个样方;分区一有8个样方、分区二有15个样方、分区三共有14个样方、分区四有11个样方、分区五包括24个样方(图1)。
对布置的样方进行定点调查,记录每个样方中的物种数、冠幅、多度和环境变量(经纬度、海拔、坡向、坡位)等。在观测记录物种时,个别物种只鉴定到属,植物命名及生活型确定参考《中国植物志》和《中国沙漠植物志》。
1.5数据处理方法
1.5.1采用α多样性(Shannon-Wiener指数(H′)、Pielou均匀度指数(E))分析植物分布特点
Shannon-Winner指数(H)的计算公式:
H=-∑PilnPiPi=Ni/N
Pielou均匀度指数(E)的计算公式:
E=H/HmaxHmax=lnS
式中,S为物种数,Ni为样地中第i种的个体数,N为样地中所有种的个体数之和。
1.5.2采用β多样性(Srensen index、Cody指数(βC))分析不同样地间物种的相似性和相互替代水平
Srensenindex=2J/(a+b) βC=(a+b-2J)/2
式中,J表示两样地间共有的种数,a和b为两样地各自的物种数。
3条样带β多样性的计算是沿西向东,分别计算样带内相邻样方间β多样性大小;对5个分区的β多样性计算是分别以1个分区与其他分区进行两两比较,得出分区间的β多样性大小。
1.5.3采用重要值和相对频度来评价物种生态优势度和适应性
重要值(%) = (相对密度+相对频度+相对盖度)/3
相对频度(rf) rf = ni/N
式中,ni表示i物种出现的样地数,N表示总样地数。
2结果与分析
2.13条样带α多样性特征
由表1可以看出,3条样带植物α多样性特征各不相同。绿洲边缘样带Shannon-Wiener指数和Pielou均匀度指数均最大;3条样带的Shannon-Wiener指数大小依次是:绿洲边缘样带>沙漠公路样带>沙漠腹地样带;Pielou均匀度指数也呈现同样的趋势,绿洲边缘样带的植物多样性分布的均匀性最佳,沙漠腹地样带的植物均匀性最低。

表1 3条样带的α多样性特征
2.23条样带植物β多样性特征
由图2可以看出:3条样带的植物β多样性总体趋势相同,自西向东随着经度的增加,物种替代水平(Cody指数)呈下降趋势,而物种的相似性水平(Sorenson指数)呈上升趋势。3条样带间β多样性变化的大小不同,沙漠公路样带的物种相似性高于其余两条样带,而物种替代水平总体低于沙漠腹地样带和绿洲边缘样带。沙漠腹地样带的物种相似性较其他两条样带稳定,变化波动小,物种相互替代水平极高。绿洲边缘样带的物种相似性较低,且变化波动较大,其物种替代性较高。

图2 3条样带植物的β多样性特征Fig.2 β-diversity characteristics of three transects
从物种组成上来看,3条样带中物种种类丰富,但相似性不大,3条样带共有物种数仅有3种,分别是倒披针叶虫实Corispermum lehmannianum、早熟猪毛菜Salsola praecox、对节刺Horaninowia ulicina。沙漠公路样带共有80个物种,样带中共有种18种,分别是梭梭Haloxylon ammodendron、早熟猪毛菜Salsola praecox、沙蓬Agriophyllum squarrosum、淡枝沙拐枣Calligonum leucocladum、蛇麻黄Ephedra distachya、疏齿千里光Senecio subdentatus、白茎绢蒿Seriphidium terraealbae、对节刺Horaninowia ulicina、倒披针叶虫实Corispermum lehmannianum、囊果苔草Carex physodes、翅鹤虱Lepechiniella lasiocarpa、镰荚黄耆Astragalus arpilobus、尖喙牻牛儿苗Erodium oxyrrhynchum、齿稃草Schismus arabicus、蓝刺头Echinops sphaerocephalus、硬萼软紫草Arnebia decumbens、假狼紫草Nnoea caspica、角果藜Ceratocarpus arenarius。沙漠腹地样带共有73个物种,样带中共有物种8种,分别是淡枝沙拐枣Calligonum leucocladum、倒披针叶虫实Corispermum lehmannianum、卷果涩荠Malcolmia scorpioides、狭果鹤虱Lappula semiglabra、早熟猪毛菜Salsola praecox、对节刺Horaninowia ulicina、镰荚黄耆Astragalus arpilobus、硬萼软紫草Arnebia decumbens。绿洲边缘的样带共有76个物种,样带中出现了8个共有种,分别是倒披针叶虫实Corispermum lehmannianum、齿稃草Schismus arabicus、早熟猪毛菜Salsola praecox、对节刺Horaninowia ulicina、弯曲四齿芥Tetracme recurvata、梭梭Haloxylon ammodendron、卷果涩荠Malcolmia scorpioides。除各样带内共有物种外,每条样带还存在各样带的独有物种,即此样带有分布而另两个样带没有的物种。比如沙漠公路附近样带,独有物种有碱韭Allium polyrhizum、小车前Plantago minuta、条叶庭芥Alyssum linifolium等6个物种群落;沙漠腹地样带,独有物种有尖花天芥菜Heliotropium acutiflorum、宽翅菘蓝Isatis violascens、细叶鸢尾Iris tenuifolia的物种群落;在绿洲边缘样带,独有柽柳Tamarix chinensis、心叶驼绒藜Ceratoides ewersmanniana、驼蹄瓣Zygophyllum fabago、枇杷柴Reaumuria songarica等9个物种群落,并且小果菘蓝Isatis minima、宽翅菘蓝Isatis violascens、细叶鸢尾Iris tenuifolia、小车前Plantago minuta、柽柳Tamarix chinensis的相对频数和重要值较大,其中小车前的相对频数和重要值最大,分别达到47.54%和15.96%(表2)。

表2 样带间的物种特异性分布特征
2.35个分区的植物α多样性特征
由表3可以看出,分区二的Shannon-Wiener指数和Pielou指数值均最大。由于分区五植物类型少及梭梭大面积退化区[30],Shannon-Wiener指数和Pielou指数值均最小。其余3个分区α多样性特征相差不大。

表3 5个分区的植物α多样性特征
2.45个分区植物β多样性特征
从表4可以看出,不同分区间的物种相似性和替代水平并不一致。分区一至分区四的物种相似性和替代性特征差异不大。分区五与其他4个分区之间物种相似性均较低,而物种相互替代水平均较高。其中分区二与分区三的物种相似性水平最高,而物种替代性最差;分区四与分区五的物种相似性最低,物种替代性最强。

表4 5个分区间植物β多样性特征
2.55个分区物种特异性分布特征
5个分区间的植物物种组成存在差异。分区一至分区四的物种组成差异不大,但是分区五土壤盐渍化严重,该区域的植物种类少,植物盖度低,因此分区五与其他4个分区的物种多样性物种组成差异很大。分区一共调查有63个物种,其中10个物种在该分区各样方中均有分布,分别是狭果鹤虱Lappula semiglabra、镰荚黄耆Astragalus arpilobus、倒披针叶虫实Corispermum lehmannianum、齿稃草Schismus arabicus、早熟猪毛菜Salsola praecox、对节刺Horaninowia ulicina、梭梭Haloxylon ammodendron、卷果涩荠Malcolmia scorpioides、白茎绢蒿Seriphidium terraealbae、疏齿千里光Senecio subdentatus。分区二调查有68个物种,共有种9种,分别是梭梭Haloxylon ammodendron、倒披针叶虫实Corispermum lehmannianum、早熟猪毛菜Salsola praecox、淡枝沙拐枣Calligonum leucocladum、齿稃草Schismus arabicus、狭果鹤虱Lappula semiglabra、角果藜Ceratocarpus arenarius、白茎绢蒿Seriphidium terraealbae、对节刺Horaninowia ulicina。分区三有77个物种,有11种共有种,分别是倒披针叶虫实Corispermum lehmannianum、齿稃草Schismus arabicus、梭梭Haloxylon ammodendron、角果藜Ceratocarpus arenarius、弯曲四齿芥Tetracme recurvata、卷果涩荠Malcolmia scorpioides、狭果鹤虱Lappula semiglabra、白茎绢蒿Seriphidium terraealbae、镰荚黄耆Astragalus arpilobus、对节刺Horaninowia ulicina、早熟猪毛菜Salsola praecox。分区四有75种植物,10种共有物种,分别是齿稃草Schismus arabicus、白茎绢蒿Seriphidium terraealbae、早熟猪毛菜Salsola praecox、梭梭Haloxylon ammodendron、镰荚黄耆Astragalus arpilobus、倒披针叶虫实Corispermum lehmannianum、卷果涩荠Malcolmia scorpioides、狭果鹤虱Lappula semiglabra、弯曲四齿芥Tetracme recurvata、对节刺Horaninowia ulicina。分区五有32个物种,该分区只有1种共有种,即梭梭Haloxylon ammodendron,在该区梭梭由于受到次生盐渍化的危害,其生长状况不良,55%左右的梭梭植株均是枯死状态(不同种群,植株的死亡率分别为37.36%、49.07%、40.85%、60.87%、50.77%、71.19%、77%、28.03%、73.28%、75.78%、56.94%、56.24%、55.69%、68.36%等),幼苗更新严重不足(不同种群,幼苗的死亡率分别为100%、55.49%、46.18%、14.04%、100%、100%、74.71%、33.26%、100%、30.29%、100%、60.92%、74.41%、30.86%等)[30]。
除了共有种在各个分区的分布很广,数量丰富之外,每个分区出现独有物种,这类物种分布范围比较小,但植物资源不容忽视(表5)。
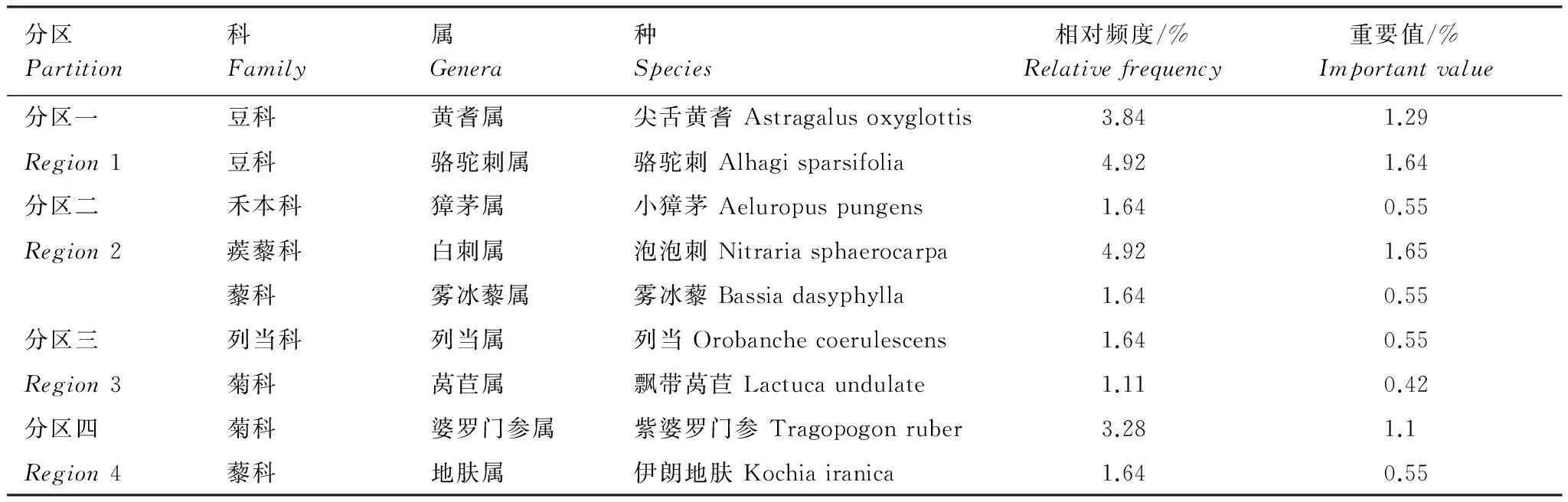
表5 分区间的物种特异性分布特征
3讨论
生物多样性分布格局与研究的尺度具有密切的关系,在不同尺度上,控制生物多样性格局的因素有较大差异,研究不同尺度的亚区域或局域的多样性特点对于全面了解区域生物多样性分布格局及其形成机制具有极大促进作用。本研究采用大样方(1000m2)密集样点的形式,对比分析了沙漠公路样带、沙漠腹地样带、绿洲边缘样带之间的多样性差异,同时根据不同生境特点,将研究区分成5个分区,全面分析古尔班通古特沙漠南部区域植物多样性分布特征。
研究发现3条样带α多样性差异较大。绿洲边缘样带位于荒漠-绿洲的过渡区,由于受到绿洲小气候等影响[31],其降水较其余两条样带多,且近十年沙漠边缘均进行了围栏保护,因此,该条样带的植物多样性和均匀性均高于其他两条样带,表明在准噶尔盆地沙漠与绿洲交错区,只要适当控制人类干扰,植被有很强的自然恢复力。沙漠公路是由于采油作业需要,在沙漠中央兴建的沥青公路,横贯整个沙漠东西端。本研究在距离公路1km以外设点,综合整个沙漠公路样带分析发现,此处植物多样性和均匀性低于绿洲边缘样带,但高于沙漠腹地样带。沙漠腹地样带位于沙漠内部,交通不便,人为干扰较低,植物多样性分布较为稳定。结合实地调查发现沙漠腹地样带低于沙漠公路样带的主要原因是沙漠腹地沙丘起伏较大,丘间面积显著少于沙漠公路周围地区,而沙漠植物多样性主体的短命植物、一年生植物和多年生草本植物等主要分布于沙丘下部和丘间[27,32]。
物种多样性在空间上有不同的变化规律[33-34]。在纬度上,物种多样性从低纬度向高纬度减少[35],由于经度上往往不存在与纬度类似的环境梯度,因而在经度上具有不同的变异规律[36-37]。古尔班通古特沙漠植物β多样性在经向上由西向东物种相似性增加,物种替代速率减弱[28]。本研究对3条样带分别分析,同样发现3条样带在经向方向上总体趋势是随着经度的增加,物种相似性增加,替代性减弱。但3条样带间β多样性变化存在差异,沙漠公路样带的物种相似性高于沙漠腹地样带和绿洲边缘样带,而物种替代水平总体低于其余两条样带,表明沙漠公路环境较为一致,物种相似高。沙漠腹地样带物种替代性高,表明在此样带沿经向方向上环境变化明显,产生相应的物种变化。绿洲边缘样带虽然受绿洲气候的强烈影响,物种多样性高,但局部环境变化较大,从而产生了物种相似性较低,变化波动较大,物种替代性居中的特点。3条样带在高经度地区物种替代性减弱,相似性增加,且趋于集中,表明高海拔下的降水增加对减少各样带样方间物种替代起到决定性,并且表现出一致的趋势。
对比发现每条样带中均有独有物种,且沙漠公路样带与绿洲边缘样带的独有种在生活型上具有明显差异。比如沙漠公路样带小果菘蓝、小车前、条叶庭芥等大部分物种属于短命植物。在绿洲边缘样带有柽柳、心叶驼绒藜、细裂补血草、驼蹄瓣、琵琶柴等多年生草本和灌木群落。而沙漠腹地样带交通困难,不易调查,通过对比发现此样带兼有沙漠公路样带与绿洲边缘样带特点,即主要是一年生和多年生共存的物种群落。
5个分区的α多样性有所不同。分区二的Shannon-Wiener指数和Pielou指数值均最大;由于分区五植物类型少及梭梭大面积退化,Shannon-Wiener指数和Pielou指数值均最小;分区一、分区三和分区四的Shannon-Wiener指数和Pielou指数值相差不大。
分区一至分区四的物种相似性和替代性特征差异不大。分区五与其他四个分区之间物种相似性均较低,而物种相互替代水平均较高。主要由于分区五位于古玛纳斯湖区,以平原为主,有2—3m的低矮沙丘,受采油等人为破坏以及地下水位下降等影响[24],土壤次生盐渍化严重,降水稀少,植被盖度低,种类很少,该区域应重视植被恢复和保护。其中分区二与分区三的物种相似性水平最高,而物种替代性最差;分区四与分区五的物种相似性最低,物种替代性最强。
同时本研究发现,除分区五以外,从其余4个分区的植物多样性物种共性来看,有7个共有种,分别是狭果鹤虱Lappula semiglabra、倒披针叶虫实Corispermum lehmannianum、齿稃草Schismus arabicus、早熟猪毛菜Salsola praecox、对节刺Horaninowia ulicina、梭梭Haloxylon ammodendron、白茎绢蒿Seriphidium terraealbae。除了梭梭和白茎绢蒿属于灌木、小灌木以外,其余5种均属于一年生草本植物,说明一年生草本植物对沙漠多样性的贡献度较高。另外,四个分区的独有物种数目较少,每个分区只有2—3种。
古尔班通古特沙漠的植物区系是古地中海退却后,第四纪形成的。黄强等对该剖面沉积物的孢粉分析表明,更新世晚期以来,古尔班通古特沙漠南部地区是灌木和草本植物生长的植被景观,藜科、蒿属和麻黄属植物是地表主要覆盖者[38- 39]。研究发现,该沙漠藜科植物是分布最广泛的物种,共调查有藜科植物14属20种,占总物种的21.7%,如倒披针叶虫实、早熟猪毛菜、对节刺、梭梭等物种的分布频度相当高,分别达到31.45%、29.21%、26.73%、18.03%。在藜科植物分布的20个物种中,研究区东部分布有17种,西部分布15种。蒿属植物在沙漠分布的主要有沙蒿和准噶尔沙蒿两种,虽两种植物在研究区东西部均有分布,但准噶尔沙蒿在东部分布更广泛。麻黄属植物在沙漠中并不常见,它主要集中在沙漠东部区域,其余地方很少甚至没有分布。本研究调查的71个属中,地中海区、西亚至中亚分布及其变型所包含属最多(28属),其次是世界分布(9属),温带分布有2属,3种分布区类型占了很大比例[27]。
Zobel等人提出的种库假说认为,一个群落的物种多样性不仅与环境条件和竞争、捕食等生态过程有关,也受区域种库限制[40]。区域种库是指一个地区可进入某一群落的潜在物种数量,它由地史过程(如冰期、地质年代)和区域过程(物种形成、迁移扩散以及消亡)所决定。按照种库假说,某一生境类型的面积越大,地质年代越古老,物种形成的机会也就越多,因而能适应和分布于该生境的物种也就越多,实际群落中的物种丰富度也就越高。种库在空间上主要有两个层次:区域种库和实际种库,前者指某一生境所拥有的潜在物种数量,主要由生物地理过程所决定;后者则为调查的群落中实际出现的物种数量,主要由竞争等生态过程和区域种库共同决定[41]。
综合本研究发现古尔班通古特沙漠南部植物多样性共同受到海盆变迁的影响,温带、世界和古地中海分布类型占有很大比例,但不同分区对比可以发现沙漠东部物种丰富度高于西部,且麻黄属植物分布广泛,应该是该沙漠西部植物扩散迁移的中心。同时也发现植物多样性与土壤质地、降水量等环境因素有极大关系[27],在本研究的3条样带之间和5个分区之间都表现的非常明显。因此古尔班通古特沙漠南部各分区间植物多样性受地史过程的影响较大,而样带之间植物多样性受土壤质地、降水量等环境因素作用。
参考文献(References):
[1]CramerMJ,WilligMR.Habitatheterogeneity,habitatassociations,androdentspeciesdiversityinaSand-Shinnery-Oaklandscape.JournalofMammalogy, 2002, 83(3): 743- 753.
[2]TewsJ,BroseU,GrimmV,TielbörgerK,WichmannMC,SchwagerM,JeltschF.Animalspeciesdiversitydrivenbyhabitatheterogeneity/diversity:theimportanceofkeystonestructures.JournalofBiogeography, 2004, 31(1): 79- 92.
[3]KadmonR,AlloucheO.Integratingtheeffectsofarea,isolation,andhabitatheterogeneityonspeciesdiversity:aunificationofislandbiogeographyandnichetheory.TheAmericanNaturalist, 2007, 170(3): 443- 454.
[4]关文彬, 冶民生, 马克明, 刘国华, 汪西林, 谭辉. 岷江干旱河谷植物群落物种周转速率与环境因子的关系. 生态学报, 2004, 24(11): 2367- 2373.
[5]RahbekC,GotelliNJ,ColwellRK,EntsmingerGL,RangelTFLVB,GravesGR.Predictingcontinental-scalepatternsofbirdspeciesrichnesswithspatiallyexplicitmodels.ProceedingsoftheRoyalSocietyB:BiologicalSciences, 2007, 274(1607): 165- 174.
[6]RapoportEH.Areography:GeographicalStrategiesofSpecies.NewYork:PergamonPress, 1982.
[7]WrightDH.Species-energytheory:Anextensionofspecies-areatheory.Oikos, 1983, 41(3): 496- 506.
[8]ColwellRK,RahbekC,GotelliNJ.Themid-domaineffectandspeciesrichnesspatterns:whathavewelearnedsofar?TheAmericanNaturalist, 2004, 163(3):E1-E23.
[9]ColwellRK,RahbekC,GotelliNJ.Themid-domaineffect:there′sababyinthebathwater.TheAmericanNaturalist, 2005, 166(5):E149-E154.
[10]RicklefsRE.Communitydiversity:relativerolesoflocalandregionalprocesses.Science, 1987, 235(4785): 167- 171.
[11]GodfrayHCJ,LawtonJH.Scaleandspeciesnumbers.TrendsinEcology&Evolution, 2001, 16(7): 400- 404.
[12]BenayasR,JoséM,ScheinerSM.Plantdiversity,biogeographyandenvironmentinIberia:patternsandpossiblecausalfactors.JournalofVegetationScience, 2002, 13(2): 245- 258.
[13]OtteD.SpeciesrichnesspatternsofNewWorlddesertgrasshoppersinrelationtoplantdiversity.JournalofBiogeography, 1976, 3(3): 197- 209.
[14]AronsonJ,ShmidaA.PlantspeciesdiversityalongaMediterranean-Desertgradientanditscorrelationtointerannualrainfallfluctuations.JournalofAridEnvironment, 1992, 23: 235- 247.
[15]AliMM,DickinsonG,MurphyKJ.PredictorsofplantdiversityinahyperariddesertwadiEcosystem.JournalofAridEnvironments, 2000, 45(3): 215- 230.
[16]BáezS,CollinsSL.ShrubinvasiondecreasesdiversityandalterscommunitystabilityinNorthernChihuahuanDesertplantcommunities.PLoSOne, 2008, 3(6):e2332.
[17]陈昌笃, 张立运, 胡文康. 古尔班通古特沙漠的沙地植物群落、区系及其分布的基本特征. 植物生态学与地植物学丛刊, 1983, 7(2): 89- 99.
[18]张立运, 陈昌笃. 论古尔班通古特沙漠植物多样性的一般特点. 生态学报, 2002, 22(11): 1923- 1932.
[19]钱亦兵, 吴兆宁, 张立运, 师庆东, 蒋进, 唐立松. 古尔班通古特沙漠生境对植物群落格局的影响. 地理学报, 2004, 59(6): 895- 902.
[20]张锦春, 王继和, 赵明, 刘虎俊, 廖空太, 徐先英. 库姆塔格沙漠南缘荒漠植物群落多样性分析. 植物生态学报, 2006, 30(3): 375- 382.
[21]吴正. 准噶尔盆地沙漠地貌发育的基本特征. 北京: 科学出版社, 1997.
[22]王雪芹, 赵从举. 古尔班通古特沙漠工程防护体系内的蚀积变化与植被的自然恢复. 干旱区地理, 2002, 25(3): 201- 207.
[23]曾晓玲, 刘彤, 张卫宾, 孙钦明, 沈雪莹, 司朗明. 古尔班通古特沙漠西部地下水位和水质变化对植被的影响. 生态学报, 2012, 32(5): 1490- 1501.
[24]钱亦兵, 吴兆宁, 张立运, 赵锐锋, 王小燕, 李有民. 古尔班通古特沙漠短命植物的空间分布特征. 科学通报, 2007, 52(19): 2299- 2306.
[25]赵怀宝, 刘彤, 雷加强, 桂东伟, 赵新俊. 古尔班通古特沙漠南部植物群落β多样性及其解释. 草业学报, 2010, 19(3): 29- 37.
[26]钱亦兵, 吴兆宁, 张立运, 赵锐锋, 王小燕, 李有民. 古尔班通古特沙漠植被与环境的关系. 生态学报, 2007, 27(7): 2802- 2811.
[27]张荣, 刘彤. 古尔班通古特沙漠南部植物多样性及群落分类. 生态学报, 2012, 32(19): 6056- 6066.
[28]张立运. 新疆莫索湾地区短命植物的初步研究. 植物生态学与地植物学丛刊, 1985, 9(3): 213- 222.
[29]刘彤, 崔运河, 翟伟, 贾亚敏, 张元杭, 谢江波. 莫索湾南缘沙漠植物群落多样性抽样方法的研究. 干旱区地理, 2006, 29(3): 367- 374.
[30]司朗明, 刘彤, 刘斌, 李磊. 古尔班通古特沙漠西部梭梭种群退化原因的对比分析. 生态学报, 2011, 31(21): 6460- 6468.
[31]冯起, 司建华, 张艳武, 姚济敏, 刘蔚, 苏永红. 极端干旱地区绿洲小气候特征及其生态意义. 地理学报, 2006, 61(1): 99- 108.
[32]刘忠权, 刘彤, 张荣, 陈辉煌. 古尔班通古特沙漠南部短命植物群落物种多样性及空间分异. 生态学杂志, 2011, 30(1): 45- 52.
[33]马克明, 傅伯杰, 周华锋. 北京东灵山地区森林的物种多样性和景观格局多样性研究. 生态学报, 1999, 19(1): 1- 7.
[34]沈泽昊, 方精云, 刘增力, 伍杰. 贡嘎山东坡植被垂直带谱的物种多样性格局分析. 植物生态学报, 2001, 25(6): 721- 732.
[35]WilligMR,KaufmanDM,StevensRD.Latitudinalgradientsofbiodiversity:pattern,process,scale,andsynthesis.AnnualReviewofEcology,Evolution,andSystematics, 2003, 34: 273- 309.
[36]BlackburnTM,GastonKJ.SpatialpatternsinthegeographicrangesizesofbirdspeciesintheNewWorld.PhilosophicalTransactionsoftheRoyalSocietyB:BiologicalSciences, 1996, 351(1342): 897- 912.
[37]LyonsSK,WilligMR.Latitudinalpatternsofrangesize:methodologicalconcernsandempiricalevaluationsforNewWorldbatsandmarsupials.Oikos, 1997, 79(3): 568- 580.
[38]陈惠中, 金炯, 董光荣. 全新世古尔班通古特沙漠演化和气候变化. 中国沙漠, 2001, 21(4): 333- 339.
[39]黄强, 周兴佳. 晚更新世晚期以来古尔班通古特沙漠南部的气候环境演化. 干旱区地理, 2000, 23(1): 55- 60.
[40]ZobelM,vanderMaarelE,DupréC.Speciespool:theconcept,itsdeterminationandsignificanceforcommunityrestoration.AppliedVegetationScience, 1998, 1(1): 55- 66.
[41]方精云, 王襄平, 唐志尧. 局域和区域过程共同控制着群落的物种多样性: 种库假说. 生物多样性, 2009, 17(6): 605-612.
基金项目:国家自然科学基金——新疆联合基金重点项目(U1130304); 国家自然科学基金资助项目(41061004)
收稿日期:2014- 02- 27; 网络出版日期:2015- 10- 10
*通讯作者
Corresponding author.E-mail: betula@126.com
DOI:10.5846/stxb201402270338
Regional differences in plant diversity in the southern Gurbantonggut desert
LUO Ning, LIU Zunchi, YU Hang, LIU Tong*
CollegeofLifeScience,ShiheziUniversity,Shihezi832000,China
Abstract:The distribution of plant diversity is influenced by the environment, such that one area is species-rich, another area species-poor and intermediate areas of moderate richness, so it is scale-dependent, so it is scale-dependent. Studying the variety and distributed regularity of plant diversity in different regions can help us understand the underlying reasons for plant community diversity and changes to this diversity, and determine how to effectively protect and use these plant resources. In this study, we focused on the south Gurbantonggut desert, a known center of plant diversity. We divided the study site along the meridional direction into the following three transects: the oasis edge, the desert highroad from Karamay to Jimusaer, and the desert hinterland. We also divided the study site into five regions, according to the height and trend of dunes and differences in habitats. We analyzed the α-diversity and β-diversity in all transects and regions, and summarized the similarities and differences in species composition. These results showed that the α-diversity in the three transects differed, with the order as follows: oasis edge transect > desert highway transect > desert hinterland transect. The β-diversity (species similarity and substitution) of the transects varied similarly with increasing longitude. According to the species composition analysis, the desert highway transect consisted mostly of ephemeral plants, the oasis edge transect had mostly perennial herbs and shrubs, and the diversity in the desert hinterland transect had a mix of ephemeral plants, perennial herbs, and shrubs. We also observed a difference in species composition among the regions. Species richness in the western part of the desert, the Mosuowan reclamation area, desert interior, and the eastern part of the desert was high, whereas that of the Manasi Lake area was relatively low. Although species similarity of the Manasi Lake was low, its species substitution rate was high, when compared with other regions. We concluded that plant diversity among the regions is driven by the geological processes of the species pool, and that plant diversity among the transects is driven by environmental factors such as soil texture and precipitation. The eastern part of the study area was determined to be the species diffusion center of the south Gurbantonggut desert.
Key Words:desert; transects; partitions; α-diversity; β-diversity; species pool hypothesis
罗宁,刘尊驰,于航,刘彤.古尔班通古特沙漠南部植物多样性的区域差异.生态学报,2016,36(12):3572- 3581.
Luo N, Liu Z C, Yu H, Liu T.Regional differences in plant diversity in the southern Gurbantonggut desert.Acta Ecologica Sinica,2016,36(12):3572- 3581.
猜你喜欢
环球时报(2022-03-29)2022-03-29 17:14:11
知识经济·中国直销(2018年7期)2018-07-27 02:49:52
东北林业大学学报(2018年7期)2018-07-20 12:14:28
散文百家·下旬刊(2016年10期)2016-11-17 21:22:22
科技资讯(2016年19期)2016-11-15 10:39:12
人间(2016年27期)2016-11-11 16:27:23
农业与技术(2016年15期)2016-11-09 07:11:20
中国水土保持科学(2016年3期)2016-07-19 03:51:24
生态学报(2016年1期)2016-03-10 06:03:14
电测与仪表(2015年8期)2015-04-09 11:50:16
