
遮阴对欧洲鹅耳枥变色期叶色及生理的影响
2018-07-14 02:33郭力宇吴驭帆祝遵凌
中南林业科技大学学报 2018年8期
郭力宇,施 曼,吴驭帆,周 琦,祝遵凌,d
(南京林业大学 a.南方现代林业协同创新中心;b.风景园林学院;c.林学院;d.艺术设计学院,江苏 南京 210037)
光照强度是影响秋色叶植物秋季叶色变化的主要环境因子之一[1],不但与植物的叶色转变程度关系密切,而且直接影响变色期的长短。同一植株外叶位较内叶位变色明显,就是由于接受光强的不同所造成[2-3]。弱光条件下叶片常表现为叶绿素含量增加,叶色加深[4],而强光对叶色有漂白作用[5]。因此,适宜的光照条件直接影响秋色叶植物的观赏价值。
欧洲鹅耳枥Carpinus betulus为桦木科Betulaceae鹅耳枥属Carpinus落叶乔木,原产于欧洲和西亚[6],树姿美丽,秋叶金黄,在园林中常以行道树或绿篱形式栽植,为著名的秋色叶树种。国内目前已在南京、上海、北京等地对其进行引种栽培,生态适应性良好[7],然而不同地区的自然环境差异显著,特别是秋季影响其变色的光照和温度因子南北差异较大,因此其在不同地区秋季叶色的呈现也有较大差异。本试验以欧洲鹅耳枥及其3个园艺品种为材料,在南京地区采用不同梯度的遮阴处理营造不同的光照强度,进而研究不同光照强度下各欧洲鹅耳枥品种叶色及其生理条件变化,以期为筛选适宜的光照强度及变色效果更好、观赏价值更高的品种提供参考,也为欧洲鹅耳枥适生地的优化筛选进一步提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为欧洲鹅耳枥Carpinus betulus及其3个园艺品种Carpinus betulus‘Beekman’、Carpinus betulus‘Frans Fontaine’、Carpinus betulus‘Lucas’,原产地法国,引入国内后盆栽管理,试验时苗龄3 a,苗高80~100 cm,规格统一。除欧洲鹅耳枥为实生苗外,其他园艺品种均为嫁接苗,砧木为欧洲鹅耳枥原种,砧木与接穗规格基本一致,常规育苗管理。以下欧洲鹅耳枥原种简称为Original,园艺品种以品种名简称。
1.2 试验设计
试验地位于南京林业大学园林试验中心(32°4′N、118°48′E),属亚热带季风气候区。2014年6月30日至2014年9月30日,选用标准透光率为50%的黑色遮阴网对各欧洲鹅耳枥进行遮阴梯度处理,双层遮阴网下透光率为25%(T1),单层遮阴网透光率为50%(T2),全光照条件下透光率为100%(TCK)为对照处理,每个处理10盆植株,每处理重复3次,共360盆植株。为避免相互遮光,每盆植株的间距为1 m。取样时间于10月20日开始,即欧洲鹅耳枥常规变色期开始前后,每隔10 d采样一次,共6次,至12月10日结束。每次取样均取树冠外围南面中上部位置基本一致的叶片进行叶色观测及生理指标的测定。
1.3 指标测定方法
通过实地拍照记录的方式进行叶色的动态观测;叶绿素与类胡萝卜素含量的测定参照张宪政[8]的方法;花色素苷含量的测定采用盐酸乙醇混合液法[9-10];营养物质的测定均参照李合生[11]的方法,可溶性糖和淀粉采用蒽酮比色法,可溶性蛋白采用考马斯亮蓝法。
1.4 数据处理
采用Excel 2016、Adobe Photoshop CS6进行数据分析与制图,采用SPSS 22.0进行方差与相关性分析。
2 结果与分析
2.1 遮阴对各欧洲鹅耳枥叶色变化的影响
叶色记录如图1所示,各欧洲鹅耳枥在同一处理水平下的变色情况基本一致,全光照条件下变色开始时间最早,变色程度最佳,由深绿色转为明黄色,其次为50%遮光条件,25%遮光条件下各品种(种)变色情况均较差。不同品种(种)变色时间存在差异。在TCK处理下,Original在11月10日叶色即出现转黄趋势,11月20日时黄色已占据大部分叶面积;Beekman、Frans Fontaine在11月20日处叶色开始转淡,11月30日时黄化明显;Lucas变色最晚,到11月30日时才呈现黄绿色,12月10日时黄色最为鲜亮,且没有褐化现象,说明Lucas在此时处于最佳观赏期。
此外,各欧洲鹅耳枥在不同处理水平下变色差异显著。随遮阴程度的加深,各欧洲鹅耳枥不同处理变色程度较相邻处理滞后10至20 d,且在T1、T2处理下叶片呈现一定暗黄、焦枯状态,这表明,遮阴不但影响叶片的显色时间和程度,同时会影响叶片的正常生理代谢,降低其秋叶的观赏价值。
2.2 遮阴对各欧洲鹅耳枥叶片色素的影响

图1 各欧洲鹅耳枥变色期叶色的动态变化Fig. 1 Dynamic changes of leaf color inCarpinus betulusin different periods of discoloration

图2 各欧洲鹅耳枥变色期叶绿素总量的动态变化Fig. 2 Dynamic changes of total chlorophyll content inCarpinus betulusin different periods of discoloration
欧洲鹅耳枥叶色转黄的过程中,叶绿素和类胡萝卜素含量的变化趋势相近。从品种(种)角度看来,叶片内叶绿素含量的变化均随时间呈现下降的趋势,但品种(种)间有一定差异(见图2)。Original和Beekman叶绿素总量均缓步下降。Frans Fontaine则在11月20日前没有显现明显的含量波动,在T1处理下小幅上升,在11月20日处达到峰值,而后迅速下降。Lucas则整体呈下降趋势。对于不同处理而言,以全光照(TCK)处理为对比参照,在11月20日处Original3个处理叶绿素总量降幅显著,降幅分别为 TCK(73.25%) > T2(39%)> T1(36%),这表明,在同一时段,光照强度的不同导致叶绿素总量降幅差异显著,从而影响欧洲鹅耳枥叶片转色程度。同样地,Beekman在11月20日处降幅为TCK(52.6%)> T1(20.7%)> T2(17%),T1、T2处理在此时段降幅均不显著,T2处理在11月30日有较大降幅,达44.4%,T1在12月10日降幅明显,达50.2%,说明随光照强度的降低,会延迟变色期欧洲鹅耳枥叶片中叶绿素总量的降低,从而延缓变色。Frans Fontaine中也呈现相近的趋势,TCK处理在11月20日时叶绿素总量降幅为39.9%,而T1、T2处理在11月30日时才出现较明显的降幅,为41.2%和35.9%,此外,T1处理在11月20日时叶绿素总量积累才达到高峰,表明重度遮阴环境下叶片仍进行叶绿素总量的累积,不利于叶绿素的分解。Lucas在整个秋冬叶片转色期,3种不同光照强度下,叶片中叶绿素总量明显高于Original、Beekman、Frans Fontaine,其在整个转色期叶绿素总量都呈不规则的波动,但整体处于稳步下降态势,没有大幅度的下降期,在11月30日时,TCK、T2、T1的降幅分别为27%、57.2%、39.5%,中度遮阴处理下反而高于全光照下的最大降幅,这代表中度遮阴的环境可能更有利于Lucas叶色的转换。
类胡萝卜素含量变化(见图3)与叶绿素总量同步,4个品种(种)3个处理的类胡萝卜素含量均随时间的推移而降低,且Original在11月20日时,类胡萝卜素降幅分别为TCK(51.3%)、T2(25.3%)、T1(26.5%),即在上述叶绿素总量大降幅时段,类胡萝卜素的降幅远比叶绿素总量的降幅小,逐渐导致了叶片由绿转黄的过程,其他3个品种也呈现了相同趋势。
各品种(种)类胡萝卜素与叶绿素的比值在不同处理下均呈现上升趋势(见图4),说明类胡萝卜素在色素中的占比增加,这是导致叶色呈现黄色的原因。TCK处理下比值大于T1、T2处理,这与叶色分析结果吻合,进一步说明全光照环境下类胡萝卜素呈色更突出。而欧洲鹅耳枥不同品种(种)间,3个处理下比值大幅变化的时间段不同,Original、Beekman在11月20日时即有显著差异,Frans Fontaine在11月30日时差异显著,Lucas差异出现的时间最晚,到12月10日才显现。再从色素占比的角度来看,类胡萝卜素与叶绿素总量的比值为0.5时,为叶片呈色的分界点,类胡萝卜素占比大于0.5时叶片开始呈现黄色,比值越大,颜色变化越明显,叶色变黄的程度也更深。Original、Frans Fontaine、Lucas在变色初期比值一直位于0.5左右,直到上述比值大幅变化时,才出现呈色的差异,Beekman的比值在变化前期则一直处于0.5以下,说明其在变色前期的绿程度最高,但在变色时间上与其他品种(种)相近。
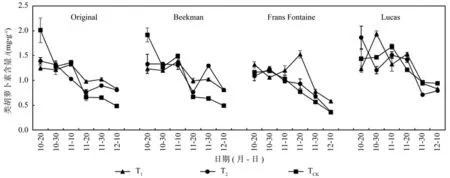
图3 各欧洲鹅耳枥变色期类胡萝卜素含量的动态变化Fig. 3 Dynamic changes of carotenoid content inCarpinus betulusin different periods of discoloration

图4 各欧洲鹅耳枥变色期类胡萝卜素与叶绿素总量比值变化Fig. 4 Changes of the ratio of carotenoids to total chlorophyll inCarpinus betulusin different periods of discoloration
各欧洲鹅耳枥花色素苷的含量动态变化随时间整体呈现下降趋势(见图5)。在Original中,T2、TCK处理下降幅为63.2%、60.8%,T1处理整体下降,但在11月10日到11月30日又进行短暂累积,随后下降,整体降幅为65.3%;Beekman品种T1、T2处理趋势相近,整体降幅为35.0%、44.9%,TCK处理下呈逐步下降趋势,降幅61.8%;T1、TCK处理下,Frans Fontaine花色素苷变化趋势为先上升后下降状态,降幅为60.4%、70.0%,T2处理下则为缓慢下降趋势,降幅为56.4%;Lucas3个处理趋势相近,均稳定下降,趋势平缓,降幅为TCK(48.0%)、T1(35.6%)、T2(29.1%)。说明花色素苷在欧洲鹅耳枥秋季叶片呈色方面作用较小。
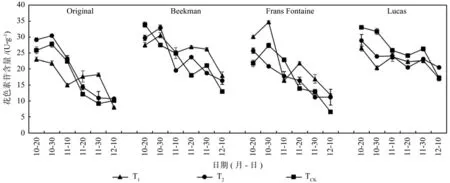
图5 各欧洲鹅耳枥变色期花色素苷含量的动态变化Fig. 5 Dynamic changes of anthocyanin content inCarpinus betulusin different periods of discoloration
2.3 遮阴对各欧洲鹅耳枥叶片生理的影响
4个品种(种)的可溶性糖含量随时间推移均呈下降的趋势(见图6),在11月10日各欧洲鹅耳枥叶片中可溶性糖含量出现上升或减缓下降的现象,随后又逐渐下降。在此期间,4个品种的可溶性糖含量均值都表现为TCK>T1>T2。可溶性淀粉含量与可溶性糖趋势接近,但与可溶性糖稍有不同,在可溶性淀粉含量(见图7)随时间下降过程中,没有出现短暂的累积趋势,而表现出稳定下降的状态,Lucas在10月20日首次采样测定时T2处理可溶性淀粉含量处于最高值,随后迅速下降,11月10日以后呈稳定状态,4个欧洲鹅耳枥品种(种)可溶性淀粉含量均值与可溶性糖同步,表现为TCK>T2>T1。

图6 各欧洲鹅耳枥变色期可溶性糖含量的动态变化Fig. 6 Dynamic changes of soluble sugar content inCarpinus betulusin different periods of discoloration

图7 各欧洲鹅耳枥变色期可溶性淀粉含量的动态变化Fig. 7 Dynamic changes of soluble starch content inCarpinus betulusin different periods of discoloration

图8 各欧洲鹅耳枥变色期可溶性蛋白含量的动态变化Fig. 8 Dynamic changes of soluble protein content inCarpinus betulusin different periods of discoloration
不同光照强度下各品种(种)欧洲鹅耳枥可溶性蛋白含量变化趋势一致(见图8),均呈先下降后上升的变化趋势,在10月30日时各处理、各品种均达到谷值,随后逐步上升。T1处理下Original涨幅(48.8%)、Beekman(54.8%)、Frans Fontaine(51.7%)、Lucas(63.6%);T2处理下 Original(54.0%)、Beekman(52.8%)、Frans Fontaine(60.6%)、Lucas(55.4%);TCK处理下Original(31.8%)、Beekman(53.1%)、Frans Fontaine(47.1%)、Lucas(53.8%)4个 品种(种)不同处理的平均值均表现为T1>T2>TCK。说明叶色变化初期,植物本身生长旺盛,随着叶色变黄,植物叶片开始进入衰老、凋落阶段,光照强度越低,越不利于其生长。
2.4 各指标间的相关性分析
全品种相关分析显示(见表1),叶绿素总量、类胡萝卜素、花色素苷、可溶性糖、可溶性淀粉、可溶性蛋白之间达到极显著水平(P<0.01),呈线性相关,其中可溶性蛋白与其他指标呈极显著线性负相关。因此在研究欧洲鹅耳枥变色期叶片的呈色时,对这些生理指标的研究具有一定的辅助意义。

表 1 各欧洲鹅耳枥生理指标的相关性分析Table 1 Correlation analysis of the physiological indexes ofCarpinus betulus
3 结论与讨论
叶片表征是肉眼对观赏性最直接的评价依据,全光照下各欧洲鹅耳枥的变色情况最佳,中度遮阴下变色情况次之,重度遮阴下变色情况最差甚至不变色。叶片表征颜色体现在叶绿素和类胡萝卜素之间的比例关系,根据色素比例的变化趋势得出结论:各欧洲鹅耳枥的叶色变换都是以类胡萝卜素含量降幅小于叶绿素总量降幅的方式实现的,从而提高了类胡萝卜素在色素总量中的占比[12],使欧洲鹅耳枥的叶片呈现黄色。同时,光照对欧洲鹅耳枥叶色的转化有着双重影响,随着遮阴度的加强,一方面降低了叶色转换的程度,另一方面延缓了叶色转换的时间,这与低光强下金叶白蜡[13]、金叶风箱果[14]等常色树种叶绿素增加、叶色返青存在区别。秋色叶植物叶片内色素含量受到生长周期、温度或光照条件等多方面因素的限制和调控,类胡萝卜素是光合合成细胞器的辅助色素,其比值的升高,可以减少光合细胞器的氧化程度。因此遮阴并不会使秋叶植物叶片出现返青的现象,但遮阴在一定程度上延缓了叶绿素的分解,从而降低了叶色转换的程度。
不同植物叶色的差异既取决于遗传因素,也受外部环境影响[15],而欧洲鹅耳枥不同品种(种)叶色的差异既受到光照影响,也来源于遗传差异。在南京地区不同欧洲鹅耳枥品种叶片色素比例转换的时间不同,这可能会导致观赏期的不同,Original、Beekman、Frans Fontaine在11月20日时,叶绿素总量降幅最大,叶片表征开始向黄色转变,而Lucas在11月30日时类胡萝卜素含量才逐渐占优,这与表观照片观测结果相近,从而可大致确定Original的观赏时期在11月20日至12月20日,Beekman和Frans Fontaine在11月30日至12月20日,Lucas观赏时间较短,在12月初至12月20日左右。原因是遮阴一定程度上延缓了色素含量的降低,但由于生长周期和温度大幅下降等因素,也会在12月中旬落叶,进而限制了秋叶的观赏期。
花色素苷是高等植物叶片中存在的一种重要的类黄酮色素,与叶绿素、类胡萝卜素共同决定叶片的呈色。当比例达到一定程度时,通常使叶片呈现红、紫、蓝色,因此花色素苷是红叶植物的主要呈色因子,而银杏[16]等黄叶植物叶片中则以类胡萝卜素呈色为主。研究中花色素苷与叶绿素、类胡萝卜素相关性达到极显著水平(P<0.01)。与榉树[17]叶色转变的研究相似,黄色单株花色素苷含量处于红绿单株之间,推测欧洲鹅耳枥秋叶变黄除受到类胡萝卜素与叶绿素比值的调控外,还与花色素苷的降解有关。从各欧洲鹅耳枥花色素苷趋势与降幅得出,不同处理中,全光照条件下花色素苷含量降幅最大,而在重度遮阴条件下,Original、Beekman、Lucas的花色苷含量上下浮动,甚至出现了短暂的累积峰,同时花色素苷与叶绿素的关联性较类胡萝卜素高,说明在欧洲鹅耳枥转色进程中,花色素苷与叶绿素随光照强度减弱可能存在协同作用。这与秋季温度降低也有一定联系,Kozlowski[18]、张开明[19]的研究表明叶绿素和花色素苷对低温的敏感性更大,从而导致类胡萝卜素掩盖了叶绿素的分解[20],实现呈色。
非结构性碳水化合物(NSC) 是植物新陈代谢过程中重要的供能物质,其动态变化反映了植物体内的碳收支状况[21],可溶性糖与淀粉是重要的反应指标。植物叶片的变色也是其对环境的响应,秋季植物落叶并转入休眠期,叶片中可溶性糖与淀粉的供应也相应减少。但在红叶植物中,糖枫[22]、枫香[23]等的研究显示,可溶性糖的含量却显著上升,这可能与可溶性糖作为花色素苷合成的前提物质有关,在红叶植物中,花色素苷是呈色的主要因子,而在欧洲鹅耳枥中,类胡萝卜素是呈色的基础,可溶性糖、花色素苷、淀粉则随时间推移呈现下降趋势,但从关联性角度而言,两者的花色素苷与可溶性糖均为正向相关。同时,3种光照条件下各欧洲鹅耳枥叶片中可溶性糖的影响趋势相近,且TCK处理可溶性糖含量均高于T1、T2,这表明:全光照条件较有利于可溶性糖和淀粉的积累,随着光照条件的减弱,植物光合作用减弱,光合产物也随即减少。可溶性淀粉趋势更加明显,均表现为TCK>T2>T1,即遮阴度越高欧洲鹅耳枥叶片NSC积累越少。处理末期可溶性糖和淀粉含量趋于稳定,各品种(种)11月10日以后的两指标的变化没有显著的上升或下降趋势,原因是植物缓慢进入休眠期,营养物质消耗减少。
可溶性蛋白也是植物体内参与生长代谢的重要内含物,本试验中可溶性蛋白含量与其他指标呈极显著的负向线性相关(P<0.01),表明欧洲鹅耳枥在秋冬季转色的过程中,植物体内的可溶性蛋白会随之发生变化,与程龙霞[24]得到的结果一致。研究中可溶性蛋白含量整体呈上升趋势,4个品种(种)含量变化幅度在TCK处理下最为缓和,10月30日时全光照、中度、重度遮阴3种处理条件下可溶性蛋白含量相当且均呈下降趋势,而随着时间的延长和遮阴强度的加重,可溶性蛋白含量显著上升,重度遮阴下的含量上升幅度最大,前人研究发现叶片中可溶性蛋白会随着温度的降低而升高。11月份温度的降低可能是导致欧洲鹅耳枥可溶性蛋白变化的主要影响因子,但在不同的遮阴度下,重度遮阴的可溶性蛋白含量在4个品种中均值最高,中度遮阴均值次之,而这2个处理下欧洲鹅耳枥叶片转色的程度低,说明光照强度的降低不但直接影响欧洲鹅耳枥叶色的转变,也对植物生长有一定的抑制作用。
本研究动态观测了4品种(种)欧洲鹅耳枥叶色转变期色素及其生理指标的变化趋势,并通过荫照处理进一步探明欧洲鹅耳枥的变色机制,但指出叶色是受多因素调控的指标,不同的光环境[25](光质、光照时长)、气候条件[26](干旱、低温)等都会对叶色的变化造成或强或弱的影响。此外,叶片内部结构也会因叶色的转变而呈现一系列的适应性变化[27],因此从多因素以及叶色对应的内部结构变化或叶色功能基因的挖掘是本研究未来的探索方向。试验结果表明:欧洲鹅耳枥变色期在全光照环境下可获得最佳的叶色表达;类胡萝卜素在色素中占比上升,叶绿素、花色素苷同步下降是欧洲鹅耳枥秋季叶色转变的直接原因,类胡萝卜素是欧洲鹅耳枥秋季叶片变黄的基础;欧洲鹅耳枥变色期NSC的积累会逐渐减少,可溶性蛋白会逐渐升高;不同欧洲鹅耳枥品种(种)秋色叶观赏期不同,在南京地区Lucas的叶色观赏期最短,Original、Beekman、Frans Fontaine的观赏期相当,在园林应用上可结合地域环境筛选适宜的品种,获取叶色最佳观赏期,实现观赏价值最大化。
猜你喜欢
语文世界(小学版)(2021年5期)2021-06-04
医药前沿(2020年36期)2020-12-03
小学生必读(低年级版)(2019年12期)2019-04-20
童话世界(2018年23期)2018-08-23
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
中南林业科技大学学报(2017年12期)2017-12-19
小学阅读指南·低年级版(2017年7期)2017-08-04
江苏农业科学(2014年3期)2014-07-16
食品工业科技(2014年6期)2014-02-27
