
3种灌木对混合盐胁迫的生理和光合响应
2018-07-14 02:33:54范文才杜丽娟郭金博殷云龙华建峰
中南林业科技大学学报 2018年8期
范文才,杜丽娟,施 钦,郭金博,殷云龙,华建峰
(1. 江苏省中国科学院植物研究所,江苏 南京 210014;2. 上海上实现代农业开发有限公司,上海 200040)
江苏省海岸线长954 km,未围垦滩涂地总面积5 001. 67 km2,占全国的28%,土壤以沙壤为主,盐分含量高,土地盐渍化现象较为严重[1]。而由于近年的围垦开发,滨海地区盐渍化及次生盐渍化面积更是急剧扩大[2],已然成为影响生态环境的严重问题,同时也是制约滨海地区植物生长的主要环境因素。盐生及耐盐植物的筛选和利用对于滨海盐碱地的生态修复具有重要意义[3],已经成为改善滨海地区土地盐渍化的重要途径[4]。
近年来,关于绿化植物耐盐性方面的研究表明,灌木由于其抗逆性强和根系较浅的特点,是滨海盐碱地快速绿化的理想植物材料[5]。王娟等[6]选择了6种常用绿化灌木,通过快相叶绿素荧光技术观测其叶片光系统Ⅱ(PSⅡ)对NaCl胁迫的快速响应特性,筛选出金银忍冬Loniceramaackii、茶条槭Acer ginnala和白丁香Syringa oblate3个耐盐灌木树种。刘昊华等[7]通过对3种滨海造林灌木树种受NaCl胁迫后叶绿素和脯氨酸等生理指标的观察分析,用隶属函数法评价3个供试树种耐盐性,由强到弱排序为:柽柳Tamarix chinensis、耐旱桑Morus alba‘Naihan’、果桑Morus alba‘Guo’。华建峰等[8]对江苏省沿海地区24种灌木与乔木进行耐盐性评价,发现灌木耐盐性普遍强于乔木,其中,海桐Pittosporum tobira、海滨木槿Hibiscus hamabo和匍地柏Sabina procumbens具有极强的耐盐性。此外,对柽柳和白刺Nitraria tangutorum等灌木耐盐机理的研究表明,盐胁迫下净光合速率、蒸腾速率等光合作用参数的变化和离子区域分布的调控能力是影响其耐盐性的重要因素,供试树种的净光合速率、蒸腾速率和气孔导度降低以响应盐胁迫,并通过茎髓部细胞内的液泡对Na+和Cl+产生区隔作用从而调控离子的区域分布以减轻盐胁迫对细胞造成的损害[9-10]。然而,目前单盐(NaCl)胁迫是对植物耐盐性生理研究的常规方法[11-13],而滨海盐渍土中的盐分多以混合盐的形式存在[14],单盐胁迫无法反映土壤中盐分组成的实际情况,并会对植物耐盐性实验结果产生一定影响。
基于江苏省滨海地区的环境特点,本实验配置了混合盐溶液,选取法青、海桐和海滨木槿3种常见灌木树种作为实验对象,研究其Na+、K+的含量及分配以及净光合速率、蒸腾速率等光合指标的响应,初步探明其耐盐机理,以期为江苏滨海盐碱地耐盐灌木材料的引种及选育提供科学依据。
1 材料与方法
1.1 试验材料
实验材料为两年生法青、海桐和海滨木槿,苗木由江苏宿迁绿杨园艺公司提供。2011年3月中旬将种苗移栽至塑料花盆中(盆高40 cm,上、下直径分别为30 cm和25 cm),每盆1株,盆底置塑料盘。栽培基质为黄土、泥炭土和珍珠岩(体积比为5∶2∶1),每盆装基质5.5 kg。苗木在室外培养2个月后,于实验开始两周前移入温室内缓苗。
1.2 试验设计
实验于2011年6月中旬在江苏省中国科学院植物研究所(35°50′ N,118°49′ E)温室内进行。混合盐溶液的配比模拟江苏省南通市如东县滨海地区盐渍土的离子组成及含量,设置3.0、5.0、8.0 g·kg-13 个盐浓度水平为处理组及 0 g·kg-1为CK组,混合盐溶液的盐分组成及含量见表1。处理组逐次加入复合盐溶液,3.0 g·kg-1加盐一次,5.0 g·kg-1处理组分两次加入,8.0 g·kg-1分 3 次,前后加盐每次间隔2 d,使其盐质量分数分别达到相应水平。实验期间定期浇水,浇水后将塑料盘中的渗出溶液重新倒入盆中,以保持各处理盐分含量,避免水分及盐分的流失。CK组及处理组的每种盐浓度水平均分别设置7个重复,随机排列。盐胁迫周期为45 d。各处理盐分组成见表1。
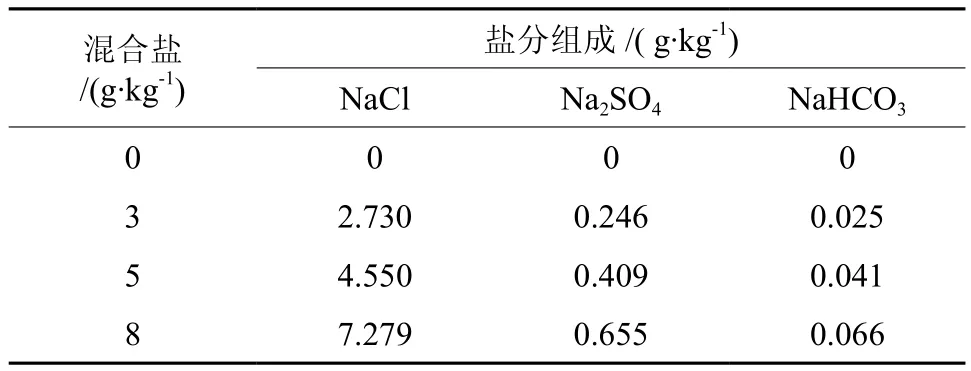
表 1 各处理的盐分组成Table 1 Salt composition of the treatments
1.3 测定指标及方法
1.3.1叶绿素(Chl)含量的测定
实验结束后,取CK组及各处理组功能叶片,擦净后用95%乙醇避光浸提样品24 h。取适量提取液分别在波长665 nm、649 nm 和470 nm 下测定吸光度,以95%乙醇作为空白对照。按照常规公式计算Chla和Chlb含量(mg/L),并计算总叶绿素含量(Chl T)=Chl a + Chl b[15]。
1.3.2Na+和K+含量的测定
取CK组及各处理组的根茎叶,烘干至恒质量后粉碎,精确称取0.75 g于50 mL规格的锥形瓶中,加入15 mL HNO3,摇匀后封口放置过夜。滴入10滴HClO4后盖上曲颈漏斗,于消煮炉上消煮 100 min(200 ℃ 40 min,300 ℃ 60 min),冷却后将消煮液定容至25 mL,样品消煮同时进行空白试验以校正方法及试剂上的误差,取10 mL定容后的消煮液定容至50 mL,于火焰光度计上测定Na+、K+。并按相同程序制备Na+、K+标准曲线[16]。
Na+(或 K+)质量分数 (%)=X·V·ts·10-4/m。 (1)式(1)中,X为标准方程求得显色液Na+(或K+)的质量浓度(μg·mL-1),V为显色液体积(mL);ts为分取倍数;m为干样质量(g)。
1.3.3光合系统参数的测定
采用美国LI-COR公司所产的Li-6400便携式光合测定仪,测定盐胁迫下3种灌木叶片的光合系统参数。测定时间段为盐胁迫处理的第43 d的9:00至11:00,使用光合测定仪内置LED红蓝光源,由其内置小钢瓶提供恒定CO2(350~450 ul·L-1),湿度控制在60%~70%。每种苗木的每种处理均随机选取4株,并随机选取其中、上部分完全展开的健康叶片3到4片进行相关测定。给定光通量密度 600 μmol·m-2s-1,停留 2 ~ 3 min 左右读数,每次测定时连续记录6~8个稳定数据。测定指标包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci),水分利用效率(WUE)为Pn和Tr的比值。
1.4 数据统计与分析
采用OriginPro 9.1制图,SPSS 19.0 进行数据统计与分析,采用新复极差法(Duncan)检验各数据组间的差异显著性,P<0.05时为差异显著。
2 结果和分析
2.1 混合盐胁迫对供试树种Na+、K+含量的影响
2.1.1供试树种Na+的含量及分配
随着混合盐胁迫浓度的增加,3个供试树种根中的Na+含量逐渐增加,且与CK均存在显著差异(P<0.05)(如图1)。盐浓度为8.0 g·kg-1时,法青、海桐和海滨木槿分别比CK增加了240%、69.67%和239%。

图1 不同浓度混合盐胁迫对3种灌木根茎叶中Na+含量的影响Fig. 1 Effects of mixed salinity stress on the Na+content in roots, stems and leaves of three different shrubs
法青茎中Na+含量随混合盐浓度的增加而显著升高(P<0.05),且在混合盐浓度为8.0 g·kg-1时含量最高,增加了690%。而海桐和海滨木槿茎中Na+含量在中、低盐浓度(盐含量<5.0 g·kg-1)下与CK差异不显著,盐浓度为8.0 g·kg-1时茎中的Na+含量显著高于CK(P<0.05),分别增加了106%和48.79%。
混合盐胁迫下法青叶片中的Na+含量在各个盐浓度处理下均显著高于CK(P<0.05),与盐浓度呈明显的正相关。海桐和海滨木槿Na+含量在3.0 g·kg-1时与CK差异不显著,5.0 g·kg-1及8.0 g·kg-1时显著性高于CK(P<0.05)。
总体来说,从不同营养器官来看,在未受到混合盐胁迫条件下,3种供试树种各器官中Na+的分布趋势均为:茎、叶差异较小,根中含量较高;而在3.0 g·kg-1~8.0 g·kg-1混合盐胁迫条件下,各树种根中Na+含量上升速率均高于茎和叶,茎叶种间差异较大,Na+含量基本为根>茎>叶。
2.1.2供试树种K+的含量及分配
与CK相比,混合盐胁迫各处理下,法青、海桐和海滨木槿根中K+含量无显著差异。混合盐胁迫后3种灌木茎中K+含量随混合盐胁迫的增强呈先增大后降低的趋势。其中,海桐和海滨木槿的K+含量均在混合盐浓度为3.0 g·kg-1时达到最高,分别是CK的1.61倍和1.26倍(P<0.05)。法青茎中K+含量则在盐浓度为5.0 g·kg-1时达到最大,为CK的1.16倍。当混合盐浓度达8.0 g·kg-1时,法青茎中K+含量显著低于CK(P<0.05),海桐和海滨木槿K+含量略低于CK,但与CK差异不显著(如图2)。

图2 不同质量分数混合盐胁迫对3种灌木根茎叶中K+含量的影响Fig.2 Effects of mixed salinity stress on the K+content in roots, stems and leaves of three different shrubs
混合盐胁迫下,各树种叶中K+含量变化差异较大。3种灌木的叶片K+含量则呈先增大后减小的变化趋势。法青和海桐叶中K+含量在混合盐浓度为3.0 g·kg-1时达到最大,但与CK差异不显著,盐浓度升高到5.0和8.0 g·kg-1时,叶中K+含量显著低于CK(P<0.05)。各个混合盐处理下海滨木槿叶中K+含量均显著高于CK(P<0.05),但各个处理间差异不显著。
总的来说,混合盐胁迫使各供试树种的K+含量下降,种间及各部位差异较大。在未受到盐胁迫情况下,3种供试树种各器官中K+的分布趋势均为:叶>根>茎。试验表明根系因最早与外界盐离子接触,因而其对K+的吸收作用在较低浓度下(3.0 g·kg-1)即被抑制,而茎、叶对K+的吸收随混合盐浓度的升高而降低。3种灌木地上部分在混合盐胁迫下积累的K+均较高,证明灌木抗逆性较强,且海滨木槿地上部分积累的K+最多,推测是其耐盐性较好的主要原因。
2.1.3混合盐胁迫对供试树种K+/Na+的影响
3个树种根中的K+/Na+均与盐胁迫浓度呈负相关,即随混合盐浓度的升高而下降。而各树种茎中的K+/Na+变化情况有所差异,法青茎中的K+/Na+随混合盐浓度的提高而逐渐降低,而海桐和海滨木槿茎中K+/Na+在3.0 g·kg-1混合盐胁迫下有所增大,分别在混合盐浓度为5.0 g·kg-1和8.0 g·kg-1时降低至CK以下。3个供试树种叶片中的K+/Na+均随混合盐胁迫浓度的增大而减小,与盐胁迫浓度呈明显的负相关。
总体来说,在未受到盐胁迫下,各树种体内保持着较高的K+/Na+,尤其是茎、叶部分。从同一植物的不同营养器官来看,法青、海桐和海滨木槿的K+/Na+均为:叶>茎>根。3个树种相比,在相同盐胁迫条件下,海滨木槿、海桐的根、茎、叶均具有更高的K+/Na+比(见表2)。

表 2 混合盐胁迫对各树种根茎叶中K+/Na+的影响†Table 2 Effects of mixed salinity stress on the K+/Na+ratio in roots, stems and leaves of three different shrubs
2.2 混合盐胁迫对供试树种叶绿素含量的影响
3.0 g·kg-1和 5.0 g·kg-1混合盐胁迫下,3 种灌木的叶绿素含量与CK相比变化不大。混合盐浓度为8.0 g·kg-1时,法青的叶绿素含量显著下降,降幅为CK的47.26%,海桐和海滨木槿叶绿素含量无明显降幅(见图3)。

图3 混合盐胁迫对3个灌木树种叶片叶绿素含量的影响Fig. 3 Effect of mixed salinity stress on leaf chlorophyll content of three different shrubs
2.3 混合盐胁迫对供试树种叶片光合系统的影响
混合盐胁迫使3种灌木的Pn下降,且各盐胁迫处理下的Pn都显著低于CK(P<0.05)。3个树种中法青的Pn变化最为剧烈。混合盐胁迫浓度为3.0 g·kg-1时,法青的Pn降幅最大,为41.98%。海桐的Pn从5.0 g·kg-1混合盐胁迫处理开始迅速下降,降幅为CK的51.77%。海滨木槿Pn变化最为平缓,在8.0 g·kg-1混合盐处理下光合速率仍能保持在较高水平。
随着混合盐浓度的增大,3种灌木的Gs和Tr逐渐减小,趋势较为一致,盐胁迫浓度越高,Gs和Tr受抑制越严重。混合盐胁迫浓度在3.0 g·kg-1时,法青的Gs和Tr变化比较剧烈,与CK相比,分别下降了71.17%和62.80%,而海桐的Gs和Tr分别下降了38.57%和41.19%,海滨木槿的Gs和Tr分别下降了53.82%和40.70%。5.0 g·kg-1浓度混合盐胁迫下,3种灌木的Gs降幅都很剧烈,法青、海桐和海滨木槿的Gs分别下降了92.20%、81.46%和85.52%,Tr分别下降了88.88%、79.29%和70.72%。方差分析显示,3种灌木的Gs和Tr在各种盐处理下与CK存在显著差异(P<0.05),而在中、高盐胁迫下(≥5.0 g·kg-1)Gs和Tr差异不显著(见图4)。
相较于上述光合参数,3种灌木在受到盐胁迫后的Ci变化趋势较为缓和。法青的Ci随着盐浓度的上升呈先下降后上升趋势,盐浓度为3.0 g·kg-1时Ci最低,降幅为CK的16.39%。海桐在盐浓度为5.0 g·kg-1时,Ci出现较大降幅,为CK的21.42%。海滨木槿的Ci受盐胁迫影响较小,5.0 g·kg-1盐胁迫时,降幅最大,仅为CK的8.40%。3种灌木受盐胁迫后均能维持较高的WUE。法青在盐浓度升高后,WUE呈先上升后下降的趋势,于盐浓度为3.0 g·kg-1时,WUE达到最高,为CK的144.06%。海桐随着盐胁迫的加重,WUE呈上升趋势,在盐浓度达到5.0 g·kg-1时上升较为明显,为CK的272.37%。海滨木槿在各盐浓度下,WUE差异不明显,较为稳定。

图4 不同质量分数混合盐胁迫对3种灌木光合特性的影响Fig. 4 Effects of mixed salinity stress on the photosynthetic characteristics of three different shrubs
3 讨论与结论
盐胁迫对植物的损害往往是通过离子毒害及渗透胁迫的方式产生作用[17]。正常生理状态下,植物细胞内各离子维持在稳定水平。受盐胁迫后,Na+含量上升,而过多的Na+会使代谢过程中的酶形成无活性蛋白从而对植物产生危害[18]。大多植物对Na+和K+的吸收具有拮抗作用,即Na+的上升一般伴随着K+的下降,而K+是植物生长发育中必须的营养元素,同时对于植物具有重要生理作用,是保证植物维持正常代谢的关键离子,因而植物在受盐胁迫后,能否维持较高水平的K+/Na+是判断其耐盐能力的重要标准[19]。杨升等[20]以K+/Na+作为重要生理指标之一科学合理的评价了卫矛Euonymus alantus、 沙 枣Elaeagnus angustifolia和美国白蜡Fraxinus Americana3 个树种的耐盐能力。莫海波等[21]对4种豆科植物的研究发现,受盐胁迫后,表现出较高K+/Na+、地上部分Na+含量较低的刺槐Robinia pseudoacacia具有更好的耐盐性。法青、海桐和海滨木槿在受混合盐胁迫后,Na+大量进入植物体内,各树种由于其根部具有较强的Na+的滞留能力,积累的Na+比茎、叶部分更多,使其地上部分Na+含量能够维持在相对较低水平以适应盐胁迫。相同盐胁迫条件下,海滨木槿和海桐的根、茎、叶相较于法青均具有更高的K+/Na+,推测3种灌木中,海滨木槿和海桐具有更好的耐盐性,这与华建峰等[8]研究结果相一致。
叶绿素含量不仅可以直观反映植物光合作用的能力,也是植物耐盐性能力评价的一个重要生理指标[22-23]。混合盐胁迫下,当盐浓度为3.0 g·kg-1时,3种灌木叶绿素含量变化不明显,当盐浓度继续增加后,海滨木槿叶绿素含量有上升趋势并在盐浓度达到8.0 g·kg-1后又轻微下降,海桐叶绿素含量整体呈下降趋势但不明显,法青叶绿素含量降幅较大。总体来看,海桐和海滨木槿在承受高浓度混合盐胁迫时叶绿素含量相较于正常水平均无明显变化,表明其对盐胁迫有较强的适应性,这与前人研究相一致[8,24]。王超宇等[25]发现在4.0和8.0 g/kg盐胁迫下,3种耐盐性较强的木本滨藜Atriplex gardneri,Atriplex tridentate,Atriplex canescens的叶绿素合成几乎没有受到影响。此外,叶绿素作为植物进行光合作用的主要色素,其含量高低与植物的净光合速率有密切关系。刘振威等[26]通过对几个品种南瓜叶片的叶绿素含量及净光合速率进行比较,证明在环境条件一致的情况下,叶绿素含量与净光合速率变化呈正相关。Pn是植物总光合速率与呼吸作用速率的差值,代表植物有机物累积速率[27],可作为衡量植物光合系统工作情况的指标[28]。本实验中,混合盐胁迫抑制了3种灌木的Pn,并且随着盐浓度的升高,3种灌木的Pn所受影响也越为严重。Farqugar[29]等认为,伴随着Gs和Ci降低的Pn降低一般由气孔限制所导致。与CK相比,除3.0 g·kg-1盐胁迫下海桐的Ci略高于CK但差异不明显外,各浓度盐胁迫下,3种灌木的Ci均有所下降,3种灌木的Gs在各盐浓度胁迫下均显著下降,表明气孔限制是混合盐胁迫下3种灌木Pn下降的原因之一,但其具体调控机理还有待深入研究。3种灌木的WUE在盐浓度为3.0 g·kg-1时均有所上升,可能是由于气孔收缩,Gs和Tr降低,从而减少了其水分的散失[30]。当盐浓度达到8.0 g·kg-1时,海桐的WUE持续上升,海滨木槿较为稳定,表明海桐和海滨木槿在盐胁迫下对水分具有较强的调节作用,而法青与低浓度盐胁迫时相比,WUE有所下降,推测其耐盐性相比海桐和海滨木槿较弱,在高浓度盐胁迫时,耐盐机制受到一定破坏。
综上研究分析,法青、海桐及海滨木槿在受混合盐胁迫后会有不同程度的生理响应。通过对这3种灌木受盐胁迫后叶绿素含量、K+/Na+及光合作用参数等重要指标的分析发现,法青在3.0 g·kg-1混合盐胁迫时,具有一定的耐盐性,而在盐浓度更高的情况下,海桐和海滨木槿表现出对盐胁迫更好的适应性。可见,海桐及海滨木槿可在沿海绿化中发挥更重要的作用,应用前景广阔。但由于本实验是在温室内展开,受土壤结构、植物生长阶段和气候差异等因素影响,具有一定局限性,若考虑将海滨木槿和海桐作为沿海防护林树种,还需进行田间实验以开展进一步研究。
猜你喜欢
农业技术与装备(2022年3期)2022-05-31 05:24:42
连云港文学(2022年2期)2022-05-10 10:43:32
青年文学家(2022年35期)2022-02-13 23:54:51
——柄果海桐
中国蜂业(2019年9期)2019-09-21 09:43:00
中老年保健(2018年3期)2018-07-12 03:26:20
散文诗(2017年17期)2018-01-31 02:34:19
作文评点报·低幼版(2016年38期)2016-11-16 09:11:02
诗林(2016年5期)2016-10-25 06:07:54
化工技术与开发(2014年1期)2014-05-09 09:29:14
未来教育家(2013年12期)2013-08-15 00:54:50
