
库尔勒香梨果实可溶性糖积累及代谢相关酶活性变化
2018-07-09 12:48艾沙江买买提张校立王继勋
新疆农业科学 2018年4期
艾沙江·买买提,张校立,梅 闯,马 凯,闫 鹏,王继勋
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测试验站,乌鲁木齐 830091)
0 引 言
【研究意义】库尔勒香梨(以下简称“香梨”)是新疆“名、特、优”水果之一,香梨已成为新疆库尔勒市农业增效,农民增收的支柱产业。但目前生产上出现香梨果肉变粗,口感多渣,石细胞含量增高,导致果实品质降低的问题。果实中糖的代谢和积累是影响果实品质的重要因素。研究果实中糖分的代谢及积累对提高香梨品质有实际意义。【前人研究进展】梨属于木本蔷薇科果树, 叶片光合产物以山梨醇为主要形态, 果实糖的积累类型属于中间转化型,果实发育早中期以积累山梨醇和淀粉为主, 至果实发育后期果糖、蔗糖和葡萄糖含量逐渐增多[1-2]。不同栽培种梨果实中糖的含量不同,果糖和葡萄糖是最主要的可溶性糖,果实中果糖含量最高, 占总糖比例的42.22%~57.02%。不同品种之间, 葡萄糖和果糖的含量相对稳定, 而山梨醇和蔗糖含量变化较大[3-9]。目前对于梨果实中糖代谢的研究,主要集中在套袋、疏花疏果等栽培措施对梨果实糖积累的影响以及酶的活性与糖积累的关系上,另外,果实糖积累过程中糖代谢关键酶的作用机制以及糖运输蛋白等方面也有一些报道[10-17]。【本研究切入点】虽然已经明确梨果实的主要糖组分及含量水平差异,但是对不同品种糖积累差异及其产生机制的研究较少,尤其相关代谢酶在库尔勒香梨果实的糖积累过程中起到作用尚不明确。研究库尔勒香梨果实发育过程中糖积累特性及相关酶活性的变化规律。【拟解决的关键问题】以库尔勒香梨为试材,分析库尔勒香梨果实发育过程中不同糖分的积累规律以及糖代谢相关酶活性的变化,研究不同发育时期果实糖分变化与酶活性之间的关系,为香梨糖分积累的分子机制研究和品质改良提供技术参数。
1 材料与方法
1.1 材 料
材料为发育过程中的库尔勒香梨(PyrusbrestschneideriRehd)果实,果实采自新疆农业科学院轮台果树资源圃15年生香梨树。选择树势健壮、管理水平一致的果树10株,于2016年4月初香梨梨花蕾期,分别在树冠的中层南部选择花蕾发育期及大小基本一致的短果枝挂牌,每短果枝保留两个果实。于花后15 d开始取样,前期每10 d取样一次,每次取大小均匀的果实30个,后期每15 d取样一次,每次取大小均匀的果实10个冷藏带回实验室,用于相关指标的测定。
1.2 方 法
参照赵世杰[18]的方法,采用葸酮比色法测定果实中可溶性糖和淀粉含量,山梨醇、葡萄糖、果糖和蔗糖的测定参考姚改芳的方法[4]采用高效液相色谱法(HPLC)。酸性转化酶(AI)、碱性转化酶(NI)、蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)的活性参照许让伟的方法[10]进行测定;山梨醇脱氢酶(SDH)和山梨醇氧化酶(SOX) 的活性测定参考Yamaguchi的方法[19]。
1.3 数据处理
所有数据采用Excel 2010和 SPSS 17.0 软件进行处理,显著水平P<0.05。采用Excel进行作图。
2 结果与分析
2.1 库尔勒香梨果实的发育动态变化规律
研究表明,香梨果实发育需要130 d左右,果实纵、横径生长曲线类似“S”型。根据果实发育期鲜重和纵横径的生长动态,香梨果实发育呈“慢-快-慢”的趋势。发育前期果实纵径生长较快,后期横径生长较快,香梨在花后70 d进入快速膨大期,坐果后75~120 d是果实快速膨大的关键阶段,至成熟时,果实增重减缓。
香梨单果重变化趋势同果实纵横径一样,在发育期呈“S”型逐渐增加的趋势,花后15~60 d为迟滞期,果实重量增加缓慢;花后51~120 d为对数增长期,果实重量增加迅速;花后 120 d以后为稳定期,果实重量增加减慢,并逐渐稳定。图1
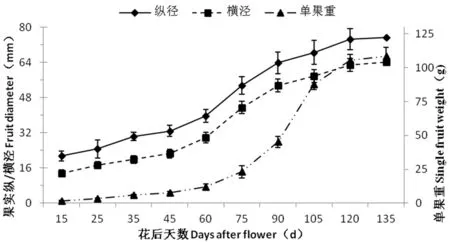
图1 库尔勒香梨果实发育过程中纵/横经、单果重变化
Fig.1 Changes of single fruit weight and fruit diameters during development of Korla fragrant pear
2.2 库尔勒香梨果实的糖积累动态变化规律
研究表明,随着果实的不断生长发育,可溶性总糖含量逐步上升,各糖组分都呈现出升高趋势。果实发育前期(花后15~60 d)果糖、葡萄糖和蔗糖含量较低并呈现出缓慢增大趋势,同时期果实中山梨醇和淀粉含量较高,其上升速度较快。花后75 d开始,香梨果实中果糖、葡萄糖和蔗糖的含量呈现出快速增长,而淀粉含量开始迅速下降,山梨醇含量的继续上升,在果实成熟期(花后105 d )山梨醇含量变化较平稳。从其构成来看,果糖为香梨果实中含量最高的可溶性糖,香梨花后135 d果实的可溶性糖为16.16%,其中果糖、山梨醇、葡萄糖和蔗糖含量分别为6.25%、3.99%、3.54%、2.35%。从糖积累最快的时期来看(花后90~135 d),花后90 d开始,香梨果实中果糖、葡萄糖和蔗糖含量快速上升, 果糖上升幅度最大,其次为葡萄糖、蔗糖的上升幅度最小,山梨醇含量却开始下降。图2

图2 库尔勒香梨果实发育过程中不同类型可溶性糖含量变化
Fig.2 Changes in different soluble sugar during fruit development in Korla fragrant pear
2.3 香梨果实发育过程中不同糖组分所占比例变化
研究表明,各组分可溶性糖在总糖中所在的比例不同。在果实发育早期,梨果实中山梨醇的含量占总糖中的比例50%以上。随着果实的发育,山梨醇所占的比例呈下降趋势,相反,果糖、葡萄糖和蔗糖所占的比例缓慢上升。果实发育早期(花后60 d以内),山梨醇的比例保持较高水平,下降速度缓慢(由51.1%降到42.1%),相应的,同时期,果糖、葡萄糖和蔗糖占总糖的比例较低,缓慢上升,果糖占总糖的比例由11.9%升至19.6%,葡萄糖占总糖的比例由8.9%升至12.2%,蔗糖占总糖的比例变化幅度很少。果实快速膨大阶段(花后70~120 d),山梨醇的比例迅速下降(由42.1%降至24.7%),果糖、葡萄糖和蔗糖所占的比例迅速上升,果实成熟期,各组分在总糖中所占的比重分别为24.7%、38.7%、22.0%、14.5%。图3

图3 香梨果实发育过程中不同糖组分所占比例变化
Fig.3 Changes in different soluble sugars ratio during fruit development in Korla fragrant pear
2.4 库尔勒香梨果实的糖积累动态变化规律
对香梨果实单果重与糖含量间相关性分析。研究表明,香梨果实大小与可溶性糖之间呈极显著的正相关,香梨果实与糖积累之间具有必然的内在联系。单果重与蔗糖、果糖、葡萄糖、山梨醇、可溶性总糖之间相关系数r分别为0.981**、0.994**、0.997**、0.860**、0.988**,均达到极显著水平。不同糖组分都与单果重高度正相关。正是由于各糖组分都与单果重高度正相关,导致各糖组分之间高度正相关。偏相关分析表明,香梨果实发育过程中,果实蔗糖含量与果糖和葡萄糖含量和极显著正相关,相关系数分别为0.836**和0.845**,与山梨醇含量显著负相关(r=-0.737*)。山梨醇是香梨果实中的重要可溶性糖,果实发育中,山梨醇含量与蔗糖、果糖和葡萄糖含量呈现负相关关系,而与总糖含量极显著正相关,说明果实发育过程中,总糖含量的增加,主要贡献是山梨醇含量的增加,果实山梨醇含量提高有利于香梨总糖的积累。表1
表1 香梨单果重与果实可溶性糖及组分相关性
Table 1 Correlation analysis of single fruit weight and soluble sugar contents of in Korla fragrant pear
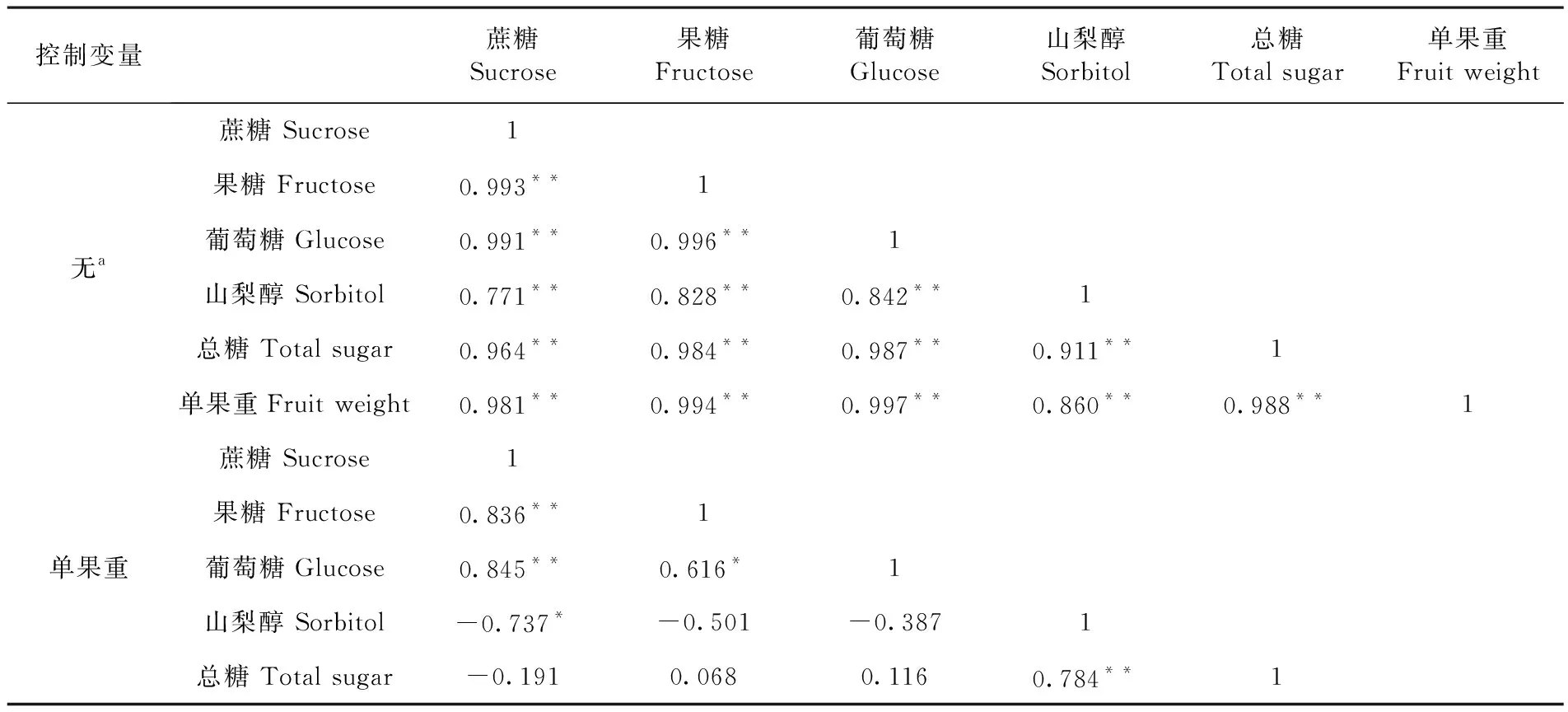
控制变量蔗糖Sucrose果糖Fructose葡萄糖Glucose山梨醇Sorbitol总糖Total sugar单果重Fruit weight无a蔗糖 Sucrose1果糖 Fructose0.993**1葡萄糖 Glucose0.991**0.996**1山梨醇 Sorbitol0.771**0.828**0.842**1总糖 Total sugar0.964**0.984**0.987**0.911**1单果重Fruit weight0.981**0.994**0.997**0.860**0.988**1单果重蔗糖 Sucrose1果糖 Fructose0.836**1葡萄糖 Glucose0.845**0.616*1山梨醇 Sorbitol-0.737*-0.501-0.3871总糖 Total sugar-0.1910.0680.1160.784**1
a. 单元格包含零阶 (Pearson) 相关,显著水平:P<0.05(*),P<0.01(**)
Significant atP<0.05(*),P<0.01(**)
2.5 香梨果实发育过程中SDH和SOX活性的变化
研究表明,在果实发育早期,香梨果实中的山梨醇脱氢酶活性均较高,随着果实的发育,NAD-SDH 活性先急剧下降,花后60~70 d出现一个低谷(分别为7.10 μmol NAD/(h·g)和7.25 μmol NAD/(h·g)),花后75 d开始逐渐升高,在果实的后期保持较高的水平。SOX的变化趋势与SDH活性趋势有类似之处,也是随着果实的发育先下降后升高。在整个果实发育阶段,香梨果实SHD和SOX酶活性变化趋势虽一致,但前者的变化幅度大,且其活性水平始终显著高于后者,这可能是香梨果实中果糖含量显著高于葡萄糖含量的原因之一。相关性分析结果表明,SDH酶活性与蔗糖含量变化显著正相关(r=0.715*),而与山梨醇和可溶性总糖含量的变化极显著负相关(r= -0.941**和r= -0.767**)。SOX酶活性只与山梨醇含量的变化极显著负相关(r= -0.777**)。表2,图4
2.6 香梨果实发育过程中SS和SPS活性变化
研究表明,果实SPS活性随果实发育逐步上升。幼果阶段(花后15~75 d)果实中SPS 活性较低且保持相对稳定,花后75 d起大幅上升,果实成熟时(花后135 d),SPS活性达到最高,为48.47 μmol/(h·g),SPS活性增高将近3倍左右。SS活性在果实生长发育初期较高,为38.03 μmol/(h·g),随后呈下降的趋势,花后60 d降到低谷(18.70 μmol/(h·g)),之后略有上升,花后75 d后SS活性又呈下降趋势,花后105 d出现第二低谷,之后随着果实的生长发育活性逐渐升高。相关性分析表明,SPS酶活性与蔗糖和果糖含量变化显著正相关(相关系数分别为r= -0.821**和r= -0.807**),而与山梨醇含量的变化极显著负相关(r= -0.838**)。SS酶活性只与可溶性总糖含量的变化显著负相关(r= -0.663**)。图5,表2

图4 库尔勒香梨果实发育过程中山梨醇脱氢酶(SDH)和山梨醇氧化酶(SOX)活性变化
Fig.4 Changes in SDH and SOX activities during the fruit development in Korla fragrant pear
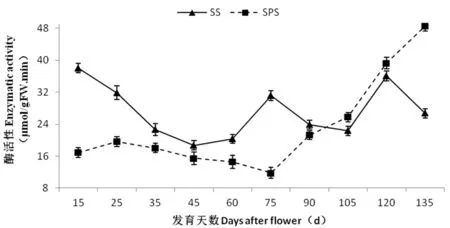
图5 库尔勒香梨果实发育过程中蔗糖合酶(SS)和蔗糖磷酸合酶(SPS)活性变化
Fig.5 Changes in SPS and SS activities during the fruit development in Korla fragrant pear
表2 香梨果实中糖组分与相关酶活性相关性
Table 2 Correlation analysis of soluble sugars and related enzymes activity in Korla fragrant pear

控制变量SDHSOXAINISPSSS无a蔗糖 Sucrose-0.030-0.0410.877**0.906**0.870**0.044果糖 Fructose-0.118-0.1110.918**0.941**0.835**0.020葡萄糖 Glucose-0.135-0.1240.924**0.944**0.805**0.017山梨醇 Sorbitol-0.619*-0.5250.923**0.863**0.413-0.256总糖 Total sugar-0.283-.0.2470.949**0.942**0.736*-0.071单果重Fruit weight-0.170-0.1550.946**0.964**0.787**-0.031单果重蔗糖 Sucrose0.715*0.579-0.817**-0.774**0.821**0.072果糖 Fructose0.4820.408-0.652-0.6230.807**-0.100葡萄糖 Glucose0.4310.391-0.735*-0.813**0.433-0.170山梨醇 Sorbitol-0.941**-0.777**0.662*0.249-0.838**-0.554总糖 Total sugar-0.767**-0.6260.276-0.261-0.437-0.663*
注:a. 单元格包含零阶 (Pearson) 相关,显著水平:P<0.05(*),P<0.01(**)
Note: Significant atP<0.05(*),P<0.01(**)
2.7 库尔勒香梨果实发育过程中AI和NI活性变化
研究表明,香梨果实中AI和NI活性变化趋势一致,且两者活性水平没有显著差异,随着果实发育而上升,pH变化均是在细胞分裂前期(花后 15~45 d)低且变化幅度较小,花后60 d起急剧上升,至花后90 d分别达到峰值5.69 μmol/(h·g)和5.50 μmol/(h·g)。在后期(从花后105 d至花后135 d)略微上升后即趋于平缓。香梨果实中的AI活性在整个发育期间约增高了4倍左右(从花后15 d的1.48 μmol/(h·g)增至花后135 d的5.85 μmol/(h·g)),NI活性在整个发育期间约增高了3倍左右(从花后15 d的2.24 μmol/(h·g)上升至花后135 d的5.78 μmol/(h·g))。相关性分析表明,AI酶活性与蔗糖和葡萄糖含量的变化显著负相关(相关系数分别为:r= -0.817**和r= -0.735*),而与山梨醇含量变化显著正相关(r=0.662*)。NI酶活性只与蔗糖和葡萄糖含量变化极显著负相关(相关系数分别为:r= -0.774**和r= -0.813*)。图6,表2
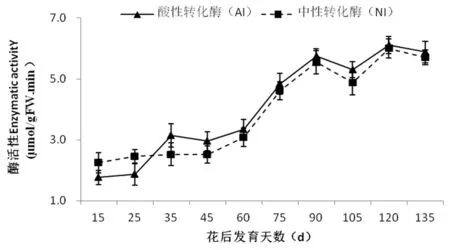
图6 库尔勒香梨果实发育过程中酸性转化酶(AI)和中性转化酶(NI)活性变化
Fig.6 Changes in AI and NI activities during the fruit development in Korla fragrant pear
3 讨 论
果实品质在很大程度上取决于所含糖的种类和数量,而其数量取决于各种糖在果实发育过程中的消长结果。不同植物果实糖的组成与积累模式不尽相同。梨等蔷薇科果树果实糖积累特征为光合产物进入果实中,在果实发育的中期同化物转换为淀粉进行积累,随后至果实发育后期同化物以糖的形式直接进入,此时,由于淀粉开始水解使果实可溶性糖含量上升[20-21]。研究发现,库尔勒香梨果实发育早中期以积累淀粉和山梨醇为主, 后期果糖、葡萄糖和蔗糖含量逐渐增多, 其中果糖含量上升最为突出并超过山梨醇成为果实中的主要糖,蔗糖含量最低,葡萄糖和山梨醇含量较接近,这与前人的研究结果基本一致[4,5,22]。果实发育早期,果糖、葡萄糖和蔗糖含量较低,可能是因为果实发育早期需要的能量和碳骨架增加,将输入的光合产物一部分用于糖酵解和三羧酸循环及随后的氨基酸和蛋白质合成,一部分转化为淀粉进行积累[23],该时期香梨果实中的淀粉含量逐渐增高,达到一个峰值;至果实发育后期, 山梨醇含量变化平缓,而其余可溶性糖含量快速增长,一部分光合产物以糖的形式直接输入, 同时淀粉也开始水解进入糖代谢, 果实中果糖、葡萄糖和蔗糖等可溶性糖含量上升。
山梨醇是梨的同化物运输的主要形式,进入果实后,大部分经过SDH和SOX 的分解作用形成葡萄糖和果糖,未被分解的积累在果实中,是影响果实品质形成的重要组分。研究中香梨果实发育90 d左右可溶性糖以山梨醇为主,说明库尔勒香梨幼果期可溶性糖以山梨醇为主,其次为果糖,葡萄糖和蔗糖含量较少;这与李馨玥等[24]在南果梨中的研究结果相同。幼果期山梨醇相关代谢酶的活性都较高这与许让伟[10]的研究结论一致,这样可以使运进的山梨醇快速地代谢以满足果实发育初期细胞分裂和生长所需要的能量和中间物。随着果实的发育SDH和SOX酶活性逐渐降低,花后60 d左右达到低谷,该时期果实中的山梨醇和淀粉含量迅速上升,淀粉含量达到最大值,而果糖和葡萄糖含量很低,变化平缓。大部分山梨醇不能转化成果糖和葡萄糖;可能该时期果实的发育需要的能量和碳骨架较少,运进的糖只有一小部分用于糖酵解和三羧酸循环及随后的氨基酸和蛋白质合成。于是果糖和葡萄糖开始在液泡中积累,淀粉合成上调,运进的糖以淀粉的形式贮存在质体中[23]。在整个果实发育阶段,香梨果实SHD和SOX酶活性变化趋势虽一致,但前者的变化幅度大,且其活性水平始终显著高于后者,以 SDH 催化山梨醇生成果糖的能力较强,对果糖积累的贡献可能相对更高[25]。这可能是香梨果实中果糖含量显著高于葡萄糖含量的原因之一。另外,淀粉开始大量降解之前,葡萄糖主要来自蔗糖的水解。山梨醇转运蛋白受到果糖和葡萄糖的竞争性抑制[26-27],为了避免山梨醇在质外体的过度积累,经过转化酶水解的蔗糖应该是少量的。由此看来,果实发育早期,蔗糖的水解对葡萄糖的积累贡献不大。试验结果表明,香梨果实发育早期葡萄糖和蔗糖含量很低,分别占总可溶性糖的8.9%和6.2%,而山梨醇的含量占总糖中的比例50%以上,该时期NI和AI活性的较低,说明葡萄糖的再生减少。果实成熟时淀粉大量的降解,葡萄糖仅仅是微量的增加,可能与该时期蔗糖的再生有关,因为果实成熟时SPS活性大增,而SPS与蔗糖的积累有显著关系。
SPS活性和蔗糖积累的趋势相一致,在蔗糖代谢中起关键作用,而SS活性对蔗糖的积累作用较小[28]。研究发现,库尔勒香梨果实发育前期蔗糖的变化幅度很少,到成熟期蔗糖含量快速上升,香梨具有在果实近成熟期积累蔗糖的特性。与其相应,果实发育前期SPS酶活性较低,随着果实的长大逐渐上升,而果实发育早期SS活性高,之后逐渐下降,到后期又上升,SPS活性和蔗糖积累的趋势相一致,这与常尚连的结论一致。香梨果实发育前期AI和NI的活性较低,随着果实发育逐渐升高,这与陈露露等[29]在对翠冠梨的研究结果相反,这可能是香梨果实中蔗糖含量始终保持较低的原因。果实将要成熟时,蔗糖含量的持续上升是和SPS酶活性上调一致。淀粉降解之前,香梨果实中的蔗糖含量保持较低水平,这一方面,蔗糖可能是果实发育前期淀粉合成与积累的来源;其次,蔗糖作为代谢调控的信号,从而调控蔗糖/淀粉代谢转化方向和强度[16, 30-34]。因此,香梨果实发育前期没有蔗糖的明显积累,可能与低含量的蔗糖仅作为单糖/多糖代谢之间的路径连接信号作用有关。果实成熟阶段蔗糖含量的增加主要是由于蔗糖的重新合成引起的。
4 结 论
库尔勒香梨果实发育早中期以积累淀粉和山梨醇为主(占可溶性总糖中的比例50%以上), 后期果糖、葡萄糖和蔗糖含量逐渐增多, 其中果糖含量上升最为突出并超过山梨醇成为果实中的主要糖(占可溶性总糖中的比例38.7%)。果实发育过程中可溶性糖的积累主要受山梨醇的影响,受蔗糖、葡萄糖和果糖积累差异的影响较小。
参考文献(References)
[1] 伍涛,陶书田,张虎平,等. 疏果对梨果实糖积累及叶片光合特性的影响[J]. 园艺学报,2011,38(11):2 041-2 048.
WU Tao,TAO Shu-tian,ZHANG Hu-ping,et al. (2011). Effects of Fruit Thinning on Fruit Sugar Accumulation and Leaf Photosynthetic Characteristics of Pear [J].ActaHorticulturaeSinica, 38(11): 2,041-2,048. (in Chinese)
[2] 黄艳花, 曾明. 梨果实糖代谢及调控因子的研究进展[J]. 植物生理学报,2013, 49(8): 709-714.
HUANG Yan-hua, ZENG Ming. (2013). Research advances in sugar metabolism and regulatory factors in pear fruits plant [J].PhysiologyJournal, 49(8): 709-714. (in Chinese)
[3] 霍月青. 砂梨品种资源糖酸及石细胞含量特点研究[D].武汉: 华中农业大学硕士学位论文,2007.
HUO Yue-qing. (2007).Studyoncharacteristicsofsugars,acidsandstonecellindifferentsandpearsfromgermplasmresources[D]. Master Dissertation: Huazhong Agricultural University, Wuhan.(in Chinese)
[4] 姚改芳,张绍铃,曹玉芬,等. 不同栽培种梨果实中可溶性糖组分及含量特征[J]. 中国农业科学,2010,43(20): 4 229-4 237.
YAO Gai-fang, ZHANG Shao-ling, CAO Yu-fen, et al. (2010). Characteristics of components and contents of soluble sugars in pear fruits from different species [J].ActaHorticulturaeSinica, 43(20): 4,229-4,237. (in Chinese)
[5] 熊碧玲,汪志辉,刘燕. 鲜黄梨果实糖积累及山梨醇转化相关酶活性的变化[J]. 中国南方果树, 2011,40(4): 24-26.
XIONG Bi-ling,WANG Zhi-hui, LIU Yan. (2011). Changes in sugar accumulation and enzymatic activities of sorbitol metabolism in developing fruit of pyrus pyrifolia cv. Sunhwang [J].SouthChinaFruits, 40 (4): 24-26. (in Chinese)
[6] 崔艳波,陈 慧,乐文全,等. 京白梨与鸭梨正反交后代果实性状遗传倾向研究[J]. 园艺学报,2011,38(2):215-224.
CUI Yan-bo,CHEN Hui,YUE Wen-quan,et al. (2011). Studies on genetic tendency of fruit characters in reciprocal crosses generation between Jingbaili and Yalipear cultivars [J].ActaHorticulturaeSinica,38(2):215-224. (in Chinese)
[7] 杨志军,乐文全,张绍铃,等. 鸭梨×京白梨杂交后代果实可溶性糖积累差异以及相关酶活性研究[J]. 园艺学报,2012,39(6):1 055-1 063.
YANG Zhi-jun,YUE Wen-quan,ZHANG Shao-ling,et al. (2012). The Difference of Soluble Sugar Accumulation and Related Enzymes Activities in Pear Fruit from Hybrid Offspring [J].ActaHorticulturaeSinica, 39(6):1,055-1,063. (in Chinese)
[8] 李芳芳,何子顺,陶书田,等. 套袋对库尔勒香梨果实发育过程中可溶性糖含量的影响[J]. 果树学报,2014,31(6): 1 072-1 078.
LI Fang-fang,HE Zi-shun,TAO Shu-tian,et al. (2014). Effects of bagging on soluble sugars contents during fruit development of Korla fragrant pear [J].JournalofFruitScience, 31(6): 1,072-1,078. (in Chinese)
[9] 王德孚,杨志军,孙江妹,等,吴俊梨不同品种果实可溶性糖积累差异及代谢相关酶活性[J]. 果树学报,2014,31(1):30-38.
WANG De -fu,YANG Zhi -jun,SUN Jiang -mei,et al. (2014). Difference in soluble sugar accumulation and related enzyme activity of fruits among different pear cultivars [J].JournalofFruitScience, 31(1): 30-38. (in Chinese)
[10] 许让伟. 砂梨果实和叶片中糖积累及代谢相关酶活性变化研究[D]. 武汉: 华中农业大学硕士论文,2009.
XU Rang-wei. (2009).Studiesonchangesofsugaraccumulatiomandrelatedenzymesactivitiesinsandpearfruitsandleaves[D]. Master Dissertation: Huazhong Agricultural University, Wuhan.(in Chinese)
[11] 于年文,李俊才,王家振,等. 南国梨果实糖积累过程中相关酶的作用[J]. 西南农业学报,2011,24(3):919-923.
YU Nian-wen,LI Jun-cai,WANG Jia-zhen,et al. (2011). Roles of related enzymes in accumulation of sugars in Nanguoli pear fruit[J].SouthwestChinaJournal, 24(3): 919-923. (in Chinese)
[12] 刘松忠,刘军,朱青青,等. 肥料种类对不同采收期黄金梨糖酸含量和风味的影响[J]. 果树学报,2012,29(5): 804-808.
LIU Song-zhong,LIU Jun,ZHU Qing-qing,et al. (2012). Effects of manure types on sugar and acid contents and flavor of pear(Pyrus pyrifolia Hawangkumbae)at different mature stages [J].JournalofFruitScience, 29(5): 804-808.(in Chinese)
[13] Li, M., Feng, F., & Cheng, L. (2012). Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development.PlosOne, 7(3): e33055.
[14] Ito, A., Sakamoto, D., & Moriguchi, T. (2012). Carbohydrate metabolism and its possible roles in endodormancy transition in japanese pear.ScientiaHorticulturae, 144(3): 187-194.
[15] 戴美松,徐飞,施泽彬,等. 砂梨山梨醇转运蛋白(SOT)基因家族成员表达特性及在果实糖积累中的作用初探[J]. 园艺学报,2015,42 (8):1 457-1 466.
DAI Mei-song,XU Fei,SHI Ze-bin,et al. (2015). Preliminary Study on Expression Characteristics of Sorbitol Transporter(SOT)Gene Family and the Role in Sugar Accumulation in Pyrus pyrifolia Fruits[J].ActaHorticulturaeSinica, 42 (8):1,457-1,466. (in Chinese)
[16] 潘俨,孟新涛,车凤斌,等. 库尔勒香梨果实发育成熟的糖代谢和呼吸代谢响应特征[J]. 中国农业科学,2016, 49(17):3 391-3 412.
PAN Yan, MENG Xin-tao, CHE Feng-bin, et al. (2016).Metabolic Profiles of Sugar Metabolism and Respiratory Metabolism of Korla Pear (Pyrus sinkiangensis Yu) Throughout Fruit Development and Ripening[J].ScientiaAgriculturaSinica, 49(17):3,391-3,412.(in Chinese)
[17] 袁晖,韦云,李馨玥,等. '南果梨'及其芽变'南红梨'果实中糖分积累与相关基因表达差异分析[J]. 果树学报, 2017,34(5):534-540.
YUAN Hui, WEI Yun, LI Xin-yue, et al. (2017). Differences in sugar accumulation and the related gene expression infruit development between'Nanguo'and its mutant'Nanhong'pears[J].JournalofFruitScience, 34(5): 534-540. (in Chinese)
[18] 赵世杰. 植物生理学实验指导[M].北京:中国农业科技出版社,1998.
ZHAO Shi-jie (1998).Experimentalguidanceofplantphysiology[M]. Beijing: China Agricultural Science and Technology Press. (in Chinese)
[19] Yamaguchi, H. (1996). Changes in the amounts of the nad-dependent sorbitol dehydrogenase and its involvement in the development of apple fruit.AmericanSocietyforHorticulturalScience, 121(5): 848-852.
[20] 王永章, 张大鹏. '红富士'苹果果实蔗糖代谢与酸性转化酶和蔗糖合酶关系的研究[J]. 园艺学报, 2001,28(3): 259-261.
WANG Yong-zhang, ZHANG Da-peng. (2001). Study on the relationships between acid invertase, sucrose synthase and sucrose metabolism in red Fuji apple fruit[J].ActaHorticulturaeSinica, 28(3): 259-261. (in Chinese)
[21] 魏建梅. 苹果(MalusdomesticaBorkh.) 果实质地品质发育及采后调控的生理和分子基础[D]. 杨凌: 西北农林科技大学博士论文,2009.
WEI Jian-mei. (2009).Studyonthephysiologicalandmelecularmechanismoffruittexturedevelopmentandpost-harvestregulationofapple(MalusdomesticaBorkh.) [D]. PhD Dissertation. Northwest A&F University, Yangling. (in Chinese)
[22] 柯凡君, 张虎平, 陶书田, 等. 套袋对梨果实发育过程中糖组分及其相关酶活性的影响[J]. 西北植物学报, 2011,31(7):1 422-1 427.
KE Fan-jun, ZHANG Hu-ping, TAO Shu-tian , et al. (2011). Sugar Component Contents and Metabol ism-related Enzyme Activities in Developing Pear Fruits after Bagging[J].ActaBot.Boreal.-Occident.Sin. ,31 (7):1,422-1,427. (in Chinese)
[23] Rolland, F., & Sheen, J. (2002). Sugar sensing and signaling in plants.PlantCell, 14(Suppl): s185.
[24] 李馨玥,李通,袁晖,等. 南果梨果实发育过程中糖分积累与相关基因表达分析[J]. 果树学报,2016,33(增刊):59-64.
LI Xin-yue,LI Tong,YUAN Hui,et al. (2016). Sugar accumulation and expression of related genes in'Nanguo'pear fruit[J].JournalofFruitScience, 33(supplement):59-64. (in Chinese)
[25] 梁东. 苹果山梨醇代谢相关基因的分子特性研究[D]. 杨凌: 西北农林科技大学博士论文,2010.
LIANG Dong (2010).Molecularcharacteristicsofgenesinvolvedinsorbitolmetabolisminapple[D] PhD Dissertation. Northwest A&F Universit, Yangling. (in Chinese)
[26] Gao, Z., Maurousset, L., Lemoine, R., Yoo, S. D., Van, N. S., & Loescher, W. (2003). Cloning, expression, and characterization of sorbitol transporters from developing sour cherry fruit and leaf sink tissues.PlantPhysiology, 131(4): 1,566-1,575.
[27] Zhang, H. P., Wu, J. Y., Tao, S. T., Wu, T., Qi, K. J., & Zhang, S. J., et al. (2014). Evidence for apoplasmic phloem unloading in pear fruit.PlantMolecularBiologyReporter, 32(4):931-939.
[28] 常尚连, 于贤昌, 于喜艳. 西瓜果实发育过程中糖分积累与相关酶活性的变化[J]. 西北农业学报,2006,15(3): 138-141.
CHANG Shang-lian, YU Xian-chang, YU Xi-yan. (2006). Change of sugar accumulation and related enzyme activity during fruit development of watermelon [J].ActaAgriculturaeBoreali-occidentalisSinica, 15(3): 138-141. (in Chinese)
[29] 陈露露,王涛,黄雪燕,等. 大棚翠冠梨果实糖积累及蔗糖代谢相关酶活性的研究[J]. 果树学报, 2011,28(3): 400-405.
CHEN Lu -lu,WANG Tao,HUANG Xue -yan,et al. (2011). Study on enzyme activities involved in sugar accumulation and sucrose metabolization in Cuiguan pear under plastic tunnel culture [J].JournalofFruitScience, 28(3): 400-405. (in Chinese)
[30] 贾海锋. 蔗糖及茉莉酸信号在草莓果实发育中的作用及其机理分析[D]. 北京: 中国农业大学博士论文,2013.
JIA Hai-feng. (2013).Rolesandmechanmismsofsucroseandjasmonicacidsignalsintheregulationofstrawberryfruitdevelopment[D]. PhD Dissertation. China Agricultural University, Beijing. (in Chinese)
[31] 陈俊伟, 张上隆, 张良诚. 果实中糖的运输、代谢与积累及其调控[J]. 植物生理与分子生物学学报, 2004,30(1): 1-10.
CHEN Jun-wei, ZHANG Shang-long, ZHANG Liang-cheng. (2004). Sugar transport, metabolism, accumulation and their regulation in fruits [J].JournalofPlantPhysiologyandMolecularBiology, 30(1): 1-10. (in Chinese)
[32] Schultz J A, Juvik J A. (2004). Current models for starch synthesis and the sugary enhancer1 (se1) mutation in Zea mays[J].PlantPhysiolBiochem, 42(6):457-464.
[33] 张红. 风味甜瓜果实酸味形成机理及糖酸遗传和积累模拟研究[D]. 杨凌: 西北农林科技大学博士论文,2009.
ZHANG Hong.(2009).MechanismoforganicacidformationandgeneticsofsugarandacidandsimulationofAccumulationinflavormelonfruit[D]. PhD Dissertation. Northwest A&F University, Yangling. (in Chinese)
[34] 郝敬虹, 李天来, 孟思达, 等, 孙利萍夜间低温对薄皮甜瓜果实糖积累及代谢相关酶活性的影响[J]. 中国农业科学, 2009,42(10): 3 592-3 599.
HAO Jing-hong, LI Tian-lai, MENG Si-da, et al. (2009). Effects of night low temperature on sugar accumulation and sugar-metabolizing enzyme activities in melon fruit [J].ScientiaAgticulturaSinica, 42(10): 3,592-3,599. (in Chinese)
猜你喜欢
中国果业信息(2022年4期)2022-11-22
作物学报(2022年1期)2022-11-05
今日农业(2022年16期)2022-09-22
河北科技师范学院学报(2022年2期)2022-08-26
农业工程学报(2022年7期)2022-07-09
石油化工(2021年9期)2021-10-18
新能源进展(2021年1期)2021-03-02
今日农业(2020年15期)2020-12-15
化工设计通讯(2020年5期)2020-01-12
今日农业(2019年11期)2019-08-15
